
2. The 6 dopaminergic systems of the brain
The brain contains a number of communication systems by means of which certain areas of the brain exchange information with each other (similar to highways within the entire road network) and which each use certain neurotransmitters.
Five of these communication systems are based on an exchange of information via dopamine (dopaminergic pathways):1
- Mesolimbic dopamine system (2.2.)
- Mesocortical dopamine system (2.3.)
- Mesostriatal (nigrostriatal) dopamine system (2.4.)
- Tuberoinfundibular dopamine system (2.5.)
- Inzertohypothalamic dopamine system (2.6.)
The cerebellum (small brain) has been recognized quite recently as containing its own dopaminergic system:
- Cerebellar dopamine system (2.7.)
There is also the dopaminergic system of the retina, through which bright daylight controls dopamine synthesis in the retina (2.8.1.), which in turn influences the circadian rhythm, as well as dopaminergic cells in the olfactory bulb (2.8.2.).
Some voices question the functional distinction between the mesocorticolimbic and nigrostriatal pathways,2 e.g. due to evidence of control of reward and aversion by the substantia nigra pars compacta, suggesting that this structure plays a role in reward, although it is not part of the mesocorticolimbic system, which is understood to be a reinforcement system.3
Within the different dopamine systems, changes in dopamine levels in individual brain regions encode different behaviors.
For example, in rats running through a maze, there is a steady increase in dopamine levels in the striatum, which peaks at the exit of the maze.4 This may encode the time estimation until the expected reward.
A high dopamine level in the (bilateral) insula, on the other hand, reduces the willingness to make an effort in order to receive rewards. The insula is said to have the function of evaluating the costs of an effort.5
2.1. Dopamine neurons in the brain
Dopamine neurons can be identified by determining their tyrosine hydroxylase immunoreactivity:6
- Mice: 21,000 to 30,000 TH-positive neurons
- Rats: 45,000 TH-positive neurons
- Primates: 160,000 to 320,000 TH-immunoreactive neurons
- human midbrain: 400,000 to 600,000 TH-positive neurons (dopamine neurons)78
- just under 600,000 for forty-year-olds
- around 350,000 for sixty-year-olds
Most dopaminergic neurons are projection neurons whose long and widely branched projections form more than a hundred thousand synapses and can therefore simultaneously influence entire cell groups in many different regions of the brain. They therefore have a very strong influence on neuronal processes and behavior.7 Human dopamine neurons of the substantia nigra are said to form over a million synapses and can have a total axon length of more than 4 m9 and therefore take several years to grow. Accordingly, brain development disorders often cause disorders in the dopaminergic system.7
Dopaminergic nerve cells are found:10
- in the ventral midbrain (predominantly), also in the ventral diencephalon
- in the telencephalon
- in the diencephalon
–> Inhibition of prolactin production- hypothalamic arcuate nucleus, A12
- subparafascicular thalamic nucleus, A11
–> Innervation of the superior olivary complex and the inferior colliculus in the brain stem, A13 (regulation of auditory processing) - medial and anterior hypothalamus, A14
- in the midbrain
- in the rostral half of the periaqueductal gray (substantia grisea periaquaeductalis, central cave gray) of the midbrain, a very small dopaminergic cell group called Aaq is found in primates7
A1512
- only exists in a few species (e.g. sheep, rodents, primates)
- is immunoreactive for tyrosine hydroxylase, a precursor of dopamine
- is located
- in ventral and dorsal components within the preoptic periventricular nucleus
- in adjacent parts of the anterior hypothalamic region
- A15 connects caudally to the dopaminergic group A14.
Specification, differentiation and maturation of dopaminergic neurons in the ventral midbrain is a complex process, influenced by various mechanisms, such as:10
- Neurulation
- Proliferation
- Differentiation of progenitor cells
- Migration
- Formation of synapses
- Formation of neuronal circuits
These mechanisms involved in the control of dopaminergic functions are controlled by external signals, such as
- Morphogens
- Growth factors
- Activation of specific gene cascades
- Activation of cellular interactions
2.2. The mesolimbic dopamine system
Part of the dopaminergic focusing, reinforcement and motivation system (sometimes also called the mesencephalic dopaminergic system or mesolimbocortical system; another part: the mesocortical system).1314
The mesolimbic, mesocortical and nigrostriatal dopamine systems consist of dopaminergic neurons with very long axons that address distant regions of the brain.15
The mesolimbic dopamine system comprises dopaminergic neurons in the ventral tegmentum (VTA) of the midbrain, in which dopamine is formed and projected to the:1617
- Nucleus accumbens in the ventral striatum
- Hippocampus (part of the limbic system)
- Amygdala (part of the limbic system)
- Septum
The limbic system controls emotional experience, its expression (pleasure / displeasure) and reward processing.
Dopamine deficiency in or injury to the nucleus accumbens results in a reduced ability to delay reward.18
Dopamine controls motor behavioral processes in the mesolimbic system in the context of reward (approaching desired objects) and reaction to novel stimuli.19
Malfunctions of the mesolimbic system:
- With ADHD:
- Problems with reinforcement mechanisms20
- Reward deferral aversion (devaluation of later rewards)16
- Delay aversion, impatience20
- Reduced frustration tolerance16
- Hyperactivity, especially in new situations2021
- Impulsiveness2021
- Disorders of behavioral inhibition/behavioral suppression20
- Changeable behavior20
- Disorder of sustained attention20
- In schizophrenia due to excess dopamine:
Activation through24
- Central stimulants
- Nicotine
- Apomorphine
- Amphetamines
- Cocaine
- Mixed inhibiting-stimulating or euphoric substances
- Alcohol
- Cannabis
- Opioids
2.3. The mesocortical dopamine system
Second part of the dopaminergic focusing, reinforcement and motivation system (sometimes also called the mesencephalic dopaminergic system; first part: the mesolimbic system).14
The mesolimbic, mesocortical and nigrostriatal dopamine systems consist of dopaminergic neurons with very long axons that address distant regions of the brain.15
It comprises connections from the VTA in Brodmann area 10 of the midbrain, where dopamine is formed, to the251416
- PFC
- The most important mesocortical brain region in ADHD
- Orbitofrontal cortex (OFC)
- Ventral cingulate gyrus
where the dopamine is released.
Dysfunctions of the mesocortical system:
- For ADHD:
- In schizophrenia due to dopamine deficiency here:
- Attention deficit disorder (positive symptom)26
- Flattening of affect (negative symptom)26
*Alogia (thought disorder with speech impoverishment, poor speech and prolonged response time)26
*Apathy = apathy, lack of excitability (non-sexual)26 - Anhedonia27
Anhedonia (inability to enjoy, reduced sense of pleasure) is also common in ADHD.
Dopamine deficiency in the mesocortical system leads to dopamine excess in the nigrostriatal system, which causes further hyperactivity and impulse problems.28
Activation through14
- Central stimulants
- Nicotine
- Apomorphine
- Amphetamines
- Cocaine
- Mixed inhibiting-stimulating or euphoric substances
- Alcohol
- Cannabis
- Opioids
Only the ADHD symptom of lack of inhibition of executive functions is caused dopaminergically by the basal ganglia (striatum, putamen), while the lack of inhibition of emotion regulation is caused noradrenergically by the hippocampus.29 Therefore, the former may be more amenable to dopaminergic treatment, while emotion regulation and affect control may be more amenable to noradrenergic treatment. This is consistent with our experience that atomoxetine optimizes emotional dysregulation in ADHD better than stimulants.
Compared to mesolimbic or nigrostriatal dopamine neurons, mesocortical DA neurons have
- higher tonic rate of fire303132
- mesoprefrontal dopamine neurons had mean discharge rates of 9.3 spikes/s and intense burst activity
- mesocingulate dopamine neurons 5.9 spikes/s and intense burst activity
- mesopiriform dopamine neurons 4.3 spikes/s and moderate burst activity
- nigrostriatal dopamine neurons 3.1 spikes/s and moderate burst activity
- more frequent burst firing333132
- higher dopamine turnover rate (2 to 4-fold)343132
- a greatly reduced responsiveness to DA agonists and antagonists3132
- a lack of tolerance to the effect of chronically administered DA antagonists3132
- selective activation through stress35, e.g. foot shocks31
- reduced development of inactivity induced by depolarization on chronic antipsychotic administration32
- Dopamine synthesis
- D2 autoreceptors
2.4. The nigrostriatal dopamine system
It involves dopaminergic neurons in the substantia nigra pars compacta that project to the basal ganglia / dorsal striatum (caudate nucleus, putamen),16 and is mainly associated with motor control36 and action selection.37
In ADHD, the dorsal striatum is the most important nigrostriatal brain region.
The mesolimbic, mesocortical and nigrostriatal dopamine systems consist of dopaminergic neurons with very long axons that address distant regions of the brain.15 Human dopamine neurons of the substantia nigra could form more than one million synapses and have a total axonal length of more than 4 m9
With their dense network, the dopaminergic neurons of the substantia nigra pars compacta “flood” the target regions in the dorsal striatum with dopamine.7 In the striatum, dopaminergic fibers form synapses every 4 μm en passant. With a half-life of around 75ms, dopamine can diffuse up to 12 μm away from its release site.38
A cascade-like spiral of projections between the VTA/substantia nigra and nucleus accumbens/striatum has been described. Dopamine neurons in the medial VTA send to medium-sized spiny projection neurons in the ventral striatum, which then project back to more lateral parts of the VTA/SNc complex, from where projections are sent to more dorsal parts of the striatum.39
Malfunctions of the nigrostriatal system:
- For ADHD:
- For Huntington’s disease:
- Hyperkinetic movement disorders42
- Tic disorders42
- In Parkinson’s disease due to dopamine deficiency or blockade of dopamine receptors by antipsychotics in this area:
- Tremor
- Rigor (muscle rigidity, muscle stiffness)
- Hypokinesia (lack of movement; slowing of movements, restricted facial expressions)
- Akinesia
2.5. The tuberoinfundibular system
The tuberoinfundibular dopamine system comprises connections from the arcuate nucleus and hypothalamus to the anterior pituitary.43
Unlike dopamine uptake in the mPFC, nucleus accumbens and caudate nucleus/putamen, which correlates with the number of dopamine receptors present, dopamine uptake in the neuroendocrine tuberoinfundibular dopamine system is lower and slower and corresponds to that of the amygdala.44
The tuberoinfundibular system and the inzertohypothalamic dopamine system have medium-length axons.45
2.5.1. Dopamine and prolactin
Dopamine inhibits the release of prolactin.
- Dopamine deficiency, e.g. due to blocked dopamine receptors in the tuberoinfundibular system, consequently increases the release of prolactin from the pituitary gland, the 2nd increment of the HPA axis.
- Prolactin has a circadian rhythm
- Maximum levels during non-REM sleep
- Major influence on sleep. (70 to 80 % of people with ADHD suffer from sleep disorders)
- Prolactin is a regulator of the emotional stress response. Prolactin is significantly increased in acute and chronic physical and psychological stress situations46 and in anxiety47.
- Conversely, a high prolactin level triggers emotional instability and anxiety.
- Prolactin is also released during orgasm.
- Prolactin increases the risk of breast cancer.46
Elevated prolactin levels (e.g. due to a lack of dopamine) cause:46
- Depressive mood / depression
- Lack of drive
- General tiredness
- States of exhaustion
- Concentration disorders
- Sleep disorders
- Mood swings
- States of anxiety
- Panic attacks
- Restlessness
- Nervousness
- Irritability
- Sensitivity to pain
- Limited social skills
- Novelty Seeking / Sensation Seeking reduced
- Changes in character
Together with the symptoms of dopamine deficiency in the mesocortical system (anhedonia = mild depression, lack of drive) and the resulting dopamine excess in the nigrostriatal system (hyperactivity, impulse control disorders), this list covers almost all of the typical ADHD symptoms.
This helps explain why dopamine-regulating stimulants are so effective in treating ADHD symptoms.
Other effects of prolactin:
Influencing homeostasis:48
- Regulation of the humoral and cellular immune response and autoimmune diseases (immunomodulation)
- Increases water transport through the breast cell membrane, sodium absorption in the small intestine.
- Promotes the formation of new blood vessels
Influence on the central nervous system:48
- Activation of dopaminergic cells
- Thereby self-regulation circle
- Stimulation of the appetite
- Anxiolytic (anxiety-relieving)
- Stress-reducing
- Regulation of oxytocin-producing nerve cells
- Stimulation of myelination in the brain
2.6. The inzertohypothalamic dopamine system
In the inzertohypothalamic dopamine system, the dopaminergic neurons are located in the hypothalamus in the catecholaminergic areas A13 and A14. These send their dopaminergic signals to the hypothalamic nuclei (PVN) and the medial preoptic area. The inzertohypothalamic dopamine system controls various functions such as nutrition, erectile function and sexual behavior.49
The tuberoinfundibular system and the inzertohypothalamic dopamine system have medium-length axons.45
Systemic administration of dopamine agonists by microinjection into the paraventricular nucleus of the hypothalamus (PVN) induces penile erection in male rats via dopamine D2 receptor activation. When microinjected into the medial preoptic area, they facilitate copulatory behavior. This is an activation of the inzertohypothalamic dopaminergic system, whose neurons originate in the catecholaminergic cell groups A13 and A14 of the hypothalamus49
2.7. The cerebellar dopamine system
The human cerebellum contains 60 to 80 % of all neurons in the brain.
ADHD is said to be particularly associated with:5051 , although the volume reduction in ADHD was just as statistically insignificant as for any other brain region52
- Posterior inferior cerebellar worm (Vermis)
- Posterior inferior lobules
Changes in the cerebellum can affect53
- visual-spatial processing
- language skills
- Agrammatism,
- Intonation changes (dysprosody)
- Syntax problems
- reduced flow of speech
- posterior regions of the cerebellum
- Executive functions53 54
- Planning
- Task organization
- Sequence of actions
- Fluency
- Working memory
- abstract thinking
- Problem-solving strategies
- Task change (task change)
- simultaneous management of several tasks
- emotional behavior53 54
- socially inappropriate behavior
- Mood disorders
- Inhibition problems
- compulsive behaviors
- Executive functions53 54
Dopaminergic midbrain nuclei transmit to the cerebellum:55
- from VTA (A10)
- from the retrorubral nucleus (A8), weaker
- from the substantia nigra pars compacta (SNpc) (A9), weaker
The cerebellum sends dopaminergic signals back to them:55
- from the vermian cerebellar cortex and from the fastigial nucleus to the ipsilateral VTA
- from the nucleus interpositus and the nucleus dentatus to the contralateral dorsal VTA and the medial and dorsal substantia nigra pars compacta
- 20 % of these fibers were bilateral
The cerebellum (here nucleus dentatus, nucleus fastigialis and nucleus interpositus posterior) is dopaminergically connected to the VTA, SNpc and nucleus caudatus55, just as the neocerebellum is dopaminergically connected to the PFC.56
In addition to the dense connections between the cerebellum and PFC, the basal ganglia and the VTA57, an independent dopaminergic network exists in the cerebellum 55 58 59 6061
Subgroups of the cerebellum show a structural specialization for cognitive functions.62
In the cerebellum, in addition to a number of other dopaminergic mechanisms, there are
- high DAT levels, especially in the posterior-inferior vermis.5559
- all dopamine receptors (DRD1 to DRD5)55
Oxygen deprivation in the cerebellum of newborn rats causes a reduction in dopamine levels and reduced expression of DRD1 and DRD2. Administration of glucose, oxygen and adrenaline partially corrects these deficiencies.63
A selective reduction of dopaminergic Purkinje neurons of the cerebellum correlated with specific impairments of cognitive functions such as behavioral flexibility, response inhibition and social recognition memory.64
Several studies have shown that the cerebellum is involved in dopaminergic neurological and psychiatric disorders such as55
- ADHD5065666768546970
- MPH also affects cerebellar function in ADHD, suggesting that behavioral changes and even motor changes in children with ADHD taking MPH could be explained by outcomes involving the cerebellum.60
- Autism715772735474757676777879
- Parkinson’s disease8081
- Schizophrenia8579
- Drug addiction8687888990
- spinocerebellar ataxia67
2.8. Other dopaminergic cells
2.8.1. Retina (A17)
Dopaminergic cells in the amacrine cell population of the retina, A17, form very short, local axons and connect only the inner and outer plexiform layers of the retina.91
A high release of dopamine in the retina adjusts vision to daylight (photopic, cone vision), while a low release adjusts vision to night light (scotopic, rod vision).92 Dopamine reduces horizontal cell coupling via the D1 receptor by phosphorylating connexins through protein kinase A, thereby closing the pore.7
This may explain the increased sensitivity to light in people with ADHD, as ADHD is associated with reduced dopamine levels,
Bright daylight controls dopamine synthesis in the retina and thus influences the circadian rhythm.
2.8.2. Olfactory bulb (A16)
The periglomerular dopamine cells of the olfactory bulb connect mitral cell dendrites in nearby neighboring glomeruli. Here, too, the axons are very short.91
2.9. Dopaminergic circadian and ultradian rhythms
Various influences control the periodicity of dopamine synthesis and the dopamine system:
- external stimuli
- Reward
- Stress
- –> temporary changes
- endogenous circadian rhythms
- –> ca, 24-hour periodic behavior of dopamine synthesis, reuptake and release in individual dopaminergic neurons
- ultradian rhythms
Ultradian rhythms have fundamentally important regulatory functions:95
- Hormone release
- Sleep phases
- Behavioral activation
- Organization of the behavioral activity
- Increasing the ability to react to environmental stimuli
In the VTA, substantia nigra and diencephalon, where dopamine is synthesized, dopamine not only influences the genetic mechanisms that control biological clocks, but is also rhythmically influenced by them, suggesting a relevant role of these brain areas in maintaining rhythmic function.96
The retina of mammals and fish is subject to a daily fluctuation in dopamine levels with a peak during the day and a decrease at night, whereby this retinal dopamine cycle is apparently influenced by melatonin.9798
Clock genes play a crucial role in neuronal functions, especially in the regulation of dopaminergic pathways. Circadian control of the dopamine system is influenced by canonical clock proteins that directly affect dopamine expression and regulation. REV-ERBalpha (nuclear receptor subfamily 1, group D, member 1, NR1D1) is a circadian nuclear receptor that negatively regulates the TTFL by targeting Bmal1 mRNA. This affects midbrain dopamine production by suppressing tyrosine hydroxylase (Th) mRNA synthesis and cholecystokinin (CCK).99100
Whether the hypothalamic nuclei are the main regulators of circadian physiology in primates is questionable. Chung et al, 2014, found little to minimal rhythmic transcript expression in nuclei typically associated with rhythmic functions, while the highest number of circadian cyclic transcripts was found in the PFC, followed by the medial globus pallidus, the putamen, the lateral globus pallidus and the mammillary bodies, which are part of the hypothalamic nuclei. In contrast, the SCN showed a very low number of cyclic transcripts. At the same time, this study demonstrated cyclic expression of dopamine D2 receptors in the putamen. However, intrinsic autonomous circadian clocks were not found in the brain areas with high cyclic gene activity.100
The expression of the clock genes is subject to rhythms that are influenced by different stimuli depending on the area of the brain:101
- in the dorsal striatum due to changes in dopamine release
- this is said to have potential effects on Parkinson’s disease and drug addiction
- in the ventral striatum due to psychological and physical stressors
- which can have an impact on severe depression
- in the limbic forebrain due to psychological and physical stressors
- which can have effects on severe depression
There is evidence of diurnal effects of dopamine reuptake inhibitors, according to which taking them at inappropriate times can cause strong dopamine peaks and subsequent dopamine drops, while taking them at appropriate times keeps dopamine levels stable for much longer. The activity of dopaminergic neurons shows intrinsic four-hour ultradian rhythms that are independent of circadian influences.95
In a mathematical modeling, the administration of dopamine reuptake inhibitors resulted in
- a sustained increase in dopamine levels during the circadian lows
- the increase was also more even, which is advantageous for medication purposes
- large fluctuations in dopamine levels during the circadian peaks
- the overall increase in extracellular dopamine was the same, but much more irregular, which is disadvantageous for medication purposes
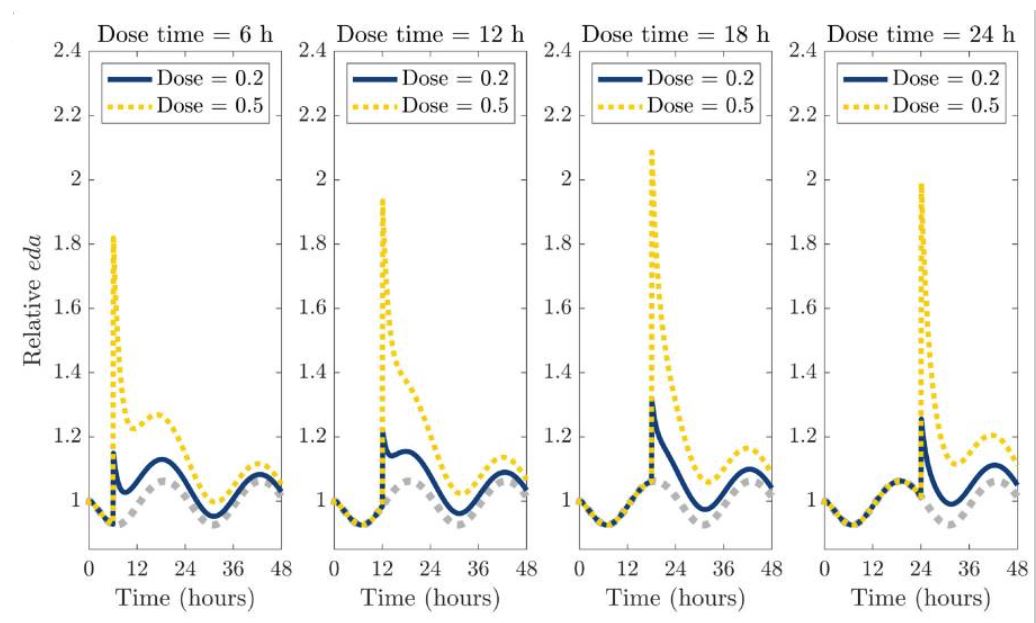
Source: The graph with the original title “Single-dose administration of a dopamine reuptake inhibitor (DRI) at different times of the day.” from and by Yao, T., & Kim, R. (2025). Mathematical modeling of dopamine rhythms and timing of dopamine reuptake inhibitors. PLoS computational biology, 21(9), e1013508. https://doi.org/10.1371/journal.pcbi.1013508 is licensed under CC BY 4.0.
The time of day when dopamine reuptake inhibitors were administered therefore also determined the duration of action.
- Modafinil administration during the circadian dopamine trough in the early morning, before the natural increase in dopamine synthesis, showed the clearest effect on alertness and performance on cognitive tasks in sleep deprivation, which also lasted for 10 hours, although the half-life of modafinil is 10 to 15 hours.102
- the increase in extracellular dopamine remained relatively low, but the circadian rhythm sets in and extracellular dopamine levels remain elevated for longer95
- A single dose of dopamine reuptake inhibitor when circadian dopamine levels are elevated and before dopamine synthesis naturally decreases due to the circadian rhythm leads to a strong but short increase in extracellular dopamine (e.g. 3 to 6 hours)95
- the diurnal effects predicted by the mathematical model are consistent with the results of clinical studies on cognitive performance
The circadian rhythm influences dopamine synthesis, dopamine release and dopamine degradation:
- Dopamine synthesis
- Dopamine release
- is subject to a circadian rhythm, which influences behavior and physiological rhythms
- however, dopamine neurons in the midbrain should not exhibit diurnal fluctuations in firing rate105
- influences melatonin106
- Melatonin release from the pineal gland
- is strongly rhythmic
- is based on the heteromerization of adrenergic receptors with dopamine D4 receptors107
- Melatonin release from the pineal gland
- is subject to a circadian rhythm, which influences behavior and physiological rhythms
- Fluctuations in dopamine levels
- also in hypothalamic neuroendocrine neurons, which control the precisely timed release of reproductive hormones such as prolactin by dopamine inhibiting prolactin
- Dopamine from tuberoinfundibular neurons is subject to a light-dark cycle activity rhythm that runs freely in constant darkness and is synchronized by light, suggesting circadian activity of tuberoinfundibular dopamine neuron activity108
- Dopamine degradation
- MAOA, which breaks down dopamine, is controlled by the internal clock and influences mood-related behaviors109
The (non-medicinal) administration of stimulants can induce a circadian rhythm in rodents, the so-called methamphetamine-sensitive circadian oscillator (MASCO). This is apparently (also) related to the dopaminergic-controlled dopamine ultradian oscillator, DUO.110 It is conceivable that the circadian sleep problems in ADHD are linked to the MASCO through dopaminergic connections.
Repeated cocaine injections show a sensitized behavioral response in wild-type mice. While this sensitization was absent in mPer1 knockout mice, mPer2 mutants showed a hypersensitized response to cocaine. Experiments on conditioned place preference revealed similar behavioral responses: mPer1 knockout mice showed no cocaine reward at all, mPer2 mutants showed a strong cocaine-induced place preference. In C57/BL6J mice, cocaine administration at different times showed that cocaine-induced behavioral sensitization and place preference are under the control of the circadian clock.111
Rensing, Koch, Rippe, Rippe (2006): Der Mensch im Stress; Psyche, Körper, Moleküle, Kapitel 4: neurobiologische Grundlagen von Stressreaktionen, Seite 89 ↥
Koevoet D, Deschamps PKH, Kenemans JL (2023): Catecholaminergic and cholinergic neuromodulation in autism spectrum disorder: A comparison to attention-deficit hyperactivity disorder. Front Neurosci. 2023 Jan 6;16:1078586. doi: 10.3389/fnins.2022.1078586. PMID: 36685234; PMCID: PMC9853424. REVIEW ↥
Ilango A, Kesner AJ, Keller KL, Stuber GD, Bonci A, Ikemoto S (2014): Similar roles of substantia nigra and ventral tegmental dopamine neurons in reward and aversion. J Neurosci. 2014 Jan 15;34(3):817-22. doi: 10.1523/JNEUROSCI.1703-13.2014. PMID: 24431440; PMCID: PMC3891961. ↥
Howe, Tierney, Sandberg, Phillips, Graybiel (2013): Prolonged Dopamine Signalling in Striatum Signals Proximity and Value of Distant Rewards; Nature. 2013 Aug 29; 500(7464): 575–579. doi: 10.1038/nature12475; PMCID: PMC3927840; NIHMSID: NIHMS507218 ↥
Treadway, Buckholtz, Cowan, Woodward, Li, Ansari, Baldwin, Schwartzman, Kessler, Zald (2012): Dopaminergic Mechanisms of Individual Differences in Human Effort-Based Decision-Making; Journal of Neuroscience 2 May 2012, 32 (18) 6170-6176; DOI: http://dx.doi.org/10.1523/JNEUROSCI.6459-11.2012 ↥
Baik (2020): Stress and the dopaminergic reward system. Exp Mol Med. 2020 Dec;52(12):1879-1890. doi: 10.1038/s12276-020-00532-4. PMID: 33257725; PMCID: PMC8080624. REVIEW ↥
Rillich (2019): Das dopaminerge System im Gehirn des Menschen: molekulare Grundlagen, Anatomie, Physiologie und Pathologie ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥
Azizi SA (2022): Monoamines: Dopamine, Norepinephrine, and Serotonin, Beyond Modulation, “Switches” That Alter the State of Target Networks. Neuroscientist. 2022 Apr;28(2):121-143. doi: 10.1177/1073858420974336. PMID: 33292070. REVIEW ↥ ↥
Pissadaki EK, Bolam JP (2013): The energy cost of action potential propagation in dopamine neurons: clues to susceptibility in Parkinson’s disease. Front Comput Neurosci. 2013 Mar 18;7:13. doi: 10.3389/fncom.2013.00013. PMID: 23515615; PMCID: PMC3600574. ↥ ↥
Speranza L, di Porzio U, Viggiano D, de Donato A, Volpicelli F (2021): Dopamine: The Neuromodulator of Long-Term Synaptic Plasticity, Reward and Movement Control. Cells. 2021 Mar 26;10(4):735. doi: 10.3390/cells10040735. PMID: 33810328; PMCID: PMC8066851. REVIEW ↥ ↥
Pignatelli A, Belluzzi O (2017): Dopaminergic Neurones in the Main Olfactory Bulb: An Overview from an Electrophysiological Perspective. Front Neuroanat. 2017 Feb 14;11:7. doi: 10.3389/fnana.2017.00007. PMID: 28261065; PMCID: PMC5306133. REVIEW ↥
van Vulpen EH, Yang CR, Nissen R, Renaud LP (1999): Hypothalamic A14 and A15 catecholamine cells provide the dopaminergic innervation to the supraoptic nucleus in rat: a combined retrograde tracer and immunohistochemical study. Neuroscience. 1999;93(2):675-80. doi: 10.1016/s0306-4522(99)00173-6. PMID: 10465451. ↥
Genro JP, Kieling C, Rohde LA, Hutz MH (2010): Attention-deficit/hyperactivity disorder and the dopaminergic hypotheses. Expert Rev Neurother. 2010 Apr;10(4):587-601. doi: 10.1586/ern.10.17. PMID: 20367210. REVIEW ↥
Edel, Vollmoeller (2006): ADHS bei Erwachsenen, Seite 110 ↥ ↥ ↥ ↥
Iversen L, Iversen F, Bloom S, Roth R (2009): Introduction to Neuropsychopharmacology, S. 211 ↥ ↥ ↥
Müller, Candrian, Kropotov (2011): ADHS – Neurodiagnostik in der Praxis, Seite 83 ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥
Gatzke-Kopp, Beauchaine (2007): Central nervous system substrates of impulsivity: Implications for the development of attention-deficit/hyperactivity disorder and conduct disorder. In: Coch, Dawson, Fischer ( Eds): Human behavior, learning, and the developing brain: Atypical development. New York: Guilford Press; 2007. pp. 239–263; 246 ↥
Cardinal, Pennicott, Sugathapala, Robbins, Everitt (2001): Impulsive choice induced in rats by lesions of the nucleus accumbens core. Science; (10.1126/science.1060818). ↥
Rensing, Koch, Rippe, Rippe (2006): Der Mensch im Stress; Psyche, Körper, Moleküle, Kapitel 4: neurobiologische Grundlagen von Stressreaktionen, Seite 90 ↥
Sagvolden, Johansen, Aase, Russell (2005): A dynamic developmental theory of attention-deficit/hyperactivity disorder (ADHD) predominantly hyperactive/impulsive and combined subtypes. Behav Brain Sci. 2005 Jun;28(3):397-419; discussion 419-68. ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥
Gatzke-Kopp, Beauchaine (2007): Central nervous system substrates of impulsivity: Implications for the development of attention-deficit/hyperactivity disorder and conduct disorder. In: Coch, Dawson, Fischer ( Eds): Human behavior, learning, and the developing brain: Atypical development. New York: Guilford Press; 2007. pp. 239–263; 253 ↥ ↥ ↥ ↥
Stahl (2000): Essential Psychopharmacology, Neuroscientific Basis and Practical Applications. Second Edition, Cambridge University Press; zitiert nach Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 66 ↥
Stahl (2000): Essential Psychopharmacology, Neuroscientific Basis and Practical Applications. Second Edition, Cambridge University Press, zitiert nach Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 66 ↥
Edel, Vollmoeller (2006): ADHS bei Erwachsenen, Seite 110, mwN ↥
Le Moal, Simon (1991): Mesocorticolimbic dopaminergic network: functional and regulatory roles; Physiological Reviews. 1 January 1991 Vol. 71 no. 1, 155-234 DOI: ↥
Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 66 ↥ ↥ ↥ ↥
Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 66 ↥
Castellanos (1997): Toward a pathophysiology of attention-deficit hyperactivity disorder. Clin. Pediatr. 36, 381-393 ↥
Müller, Candrian, Kropotov (2011): ADHS – Neurodiagnostik in der Praxis, Seite 85 ↥
Chiodo LA, Bannon MJ, Grace AA, Roth RH, Bunney BS (1984): Evidence for the absence of impulse-regulating somatodendritic and synthesis-modulating nerve terminal autoreceptors on subpopulations of mesocortical dopamine neurons. Neuroscience. 1984 May;12(1):1-16. doi: 10.1016/0306-4522(84)90133-7. PMID: 6462443. ↥
Bannon MJ, Roth RH (1983): Pharmacology of mesocortical dopamine neurons. Pharmacol Rev. 1983 Mar;35(1):53-68. PMID: 6138783. REVIEW ↥ ↥ ↥ ↥ ↥ ↥
Iversen L, Iversen F, Bloom S, Roth R (2009): Introduction to Neuropsychopharmacology, S. 208 ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥
White FJ, Wang RY (1983): Comparison of the effects of chronic haloperidol treatment on A9 and A10 dopamine neurons in the rat. Life Sci. 1983 Feb 28;32(9):983-93. doi: 10.1016/0024-3205(83)90929-3. PMID: 6827927. ↥
Bannon MJ, Bunney EB, Roth RH (1981): Mesocortical dopamine neurons: rapid transmitter turnover compared to other brain catecholamine systems. Brain Res. 1981 Aug 10;218(1-2):376-82. PMID: 7272743. ↥
Finlay, Zigmond (1997): The effects of stress on central dopaminergic neurons: possible clinical implications. Neurochem Res. 1997 Nov;22(11):1387-94. REVIEW ↥ ↥ ↥
Liu, Kaeser (2019): Mechanisms and regulation of dopamine release. Curr Opin Neurobiol. 2019 Aug;57:46-53. doi: 10.1016/j.conb.2019.01.001. PMID: 30769276; PMCID: PMC6629510. ↥
Liu, Goel, Kaeser (2021): Spatial and temporal scales of dopamine transmission. Nat Rev Neurosci. 2021 Jun;22(6):345-358. doi: 10.1038/s41583-021-00455-7. PMID: 33837376; PMCID: PMC8220193. ↥
Seamans JK, Yang CR (2004): The principal features and mechanisms of dopamine modulation in the prefrontal cortex. Prog Neurobiol. 2004 Sep;74(1):1-58. doi: 10.1016/j.pneurobio.2004.05.006. Erratum in: Prog Neurobiol. 2004 Dec;74(5):321. PMID: 15381316. ↥
Morikawa H, Paladini CA (2011): Dynamic regulation of midbrain dopamine neuron activity: intrinsic, synaptic, and plasticity mechanisms. Neuroscience. 2011 Dec 15;198:95-111. doi: 10.1016/j.neuroscience.2011.08.023. PMID: 21872647; PMCID: PMC3221882. REVIEW ↥
Castellanos 1997, zitiert nach Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 67 ↥ ↥
Gatzke-Kopp, Beauchaine (2007): Central nervous system substrates of impulsivity: Implications for the development of attention-deficit/hyperactivity disorder and conduct disorder. In: Coch, Dawson, Fischer (Eds): Human behavior, learning, and the developing brain: Atypical development. New York: Guilford Press; 2007. pp. 239–263; 253 ↥ ↥
Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 67 ↥ ↥
Jayanti S, Dalla Verde C, Tiribelli C, Gazzin S (2023): Inflammation, Dopaminergic Brain and Bilirubin. Int J Mol Sci. 2023 Jul 14;24(14):11478. doi: 10.3390/ijms241411478. PMID: 37511235; PMCID: PMC10380707. ↥
Garris PA, Wightman RM (1994): Different kinetics govern dopaminergic transmission in the amygdala, prefrontal cortex, and striatum: an in vivo voltammetric study. J Neurosci. 1994 Jan;14(1):442-50. doi: 10.1523/JNEUROSCI.14-01-00442.1994. PMID: 8283249; PMCID: PMC6576851. ↥
Iversen L, Iversen F, Bloom S, Roth R (2009): Introduction to Neuropsychopharmacology, S. 212 ↥ ↥
Prolaktin: Wikipedia, Stand 04.07.2016 ↥ ↥ ↥
Athanasoulia (2011): Prolaktinome und Psyche, Max Planck Institut für Psychatrie ↥
Melis, Sanna, Argiolas (2022): Dopamine, Erectile Function and Male Sexual Behavior from the Past to the Present: A Review. Brain Sci. 2022 Jun 24;12(7):826. doi: 10.3390/brainsci12070826. PMID: 35884633; PMCID: PMC9312911. REVIEW ↥ ↥
Braga-Neto P, Barbosa Dos Santos MW, Scott SSO, Munhoz RP, Novis LE, Pedroso JL, Barsottini OGP (2025): The cerebellum and psychiatric disorders: unraveling its role in mental health. Arq Neuropsiquiatr. 2025 Aug;83(8):1-8. doi: 10.1055/s-0045-1810408. PMID: 40886707. REVIEW ↥ ↥
Cortese S (2012): The neurobiology and genetics of Attention-Deficit/Hyperactivity Disorder (ADHD): what every clinician should know. Eur J Paediatr Neurol. 2012 Sep;16(5):422-33. doi: 10.1016/j.ejpn.2012.01.009. PMID: 22306277. REVIEW ↥
Valera EM, Faraone SV, Murray KE, Seidman LJ (2007): Meta-analysis of structural imaging findings in attention-deficit/hyperactivity disorder. Biol Psychiatry. 2007 Jun 15;61(12):1361-9. doi: 10.1016/j.biopsych.2006.06.011. PMID: 16950217. METASTUDY ↥
Jacobi H, Faber J, Timmann D, Klockgether T (2021): Update cerebellum and cognition. J Neurol. 2021 Oct;268(10):3921-3925. doi: 10.1007/s00415-021-10486-w. PMID: 33656586; PMCID: PMC8463403. ↥ ↥ ↥
Bruchhage MMK, Bucci MP, Becker EBE (2018): Cerebellar involvement in autism and ADHD. Handb Clin Neurol. 2018;155:61-72. doi: 10.1016/B978-0-444-64189-2.00004-4. PMID: 29891077. REVIEW ↥ ↥ ↥ ↥
Flace P, Livrea P, Basile GA, Galletta D, Bizzoca A, Gennarini G, Bertino S, Branca JJV, Gulisano M, Bianconi S, Bramanti A, Anastasi G (2021): The Cerebellar Dopaminergic System. Front Syst Neurosci. 2021 Aug 5;15:650614. doi: 10.3389/fnsys.2021.650614. PMID: 34421548; PMCID: PMC8375553. REVIEW ↥ ↥ ↥ ↥ ↥ ↥ ↥
Habas C (2021): Functional Connectivity of the Cognitive Cerebellum. Front Syst Neurosci. 2021 Apr 8;15:642225. doi: 10.3389/fnsys.2021.642225. PMID: 33897382; PMCID: PMC8060696. REVIEW ↥
Zhou P, Peng S, Wen S, Lan Q, Zhuang Y, Li X, Shi M, Zhang C (2024): The Cerebellum-Ventral Tegmental Area Microcircuit and Its Implications for Autism Spectrum Disorder: A Narrative Review. Neuropsychiatr Dis Treat. 2024 Oct 29;20:2039-2048. doi: 10.2147/NDT.S485487. PMID: 39494383; PMCID: PMC11531233. REVIEW ↥ ↥
Li C, Saliba NB, Martin H, Losurdo NA, Kolahdouzan K, Siddiqui R, Medeiros D, Li W (2023): Purkinje cell dopaminergic inputs to astrocytes regulate cerebellar-dependent behavior. Nat Commun. 2023 Mar 23;14(1):1613. doi: 10.1038/s41467-023-37319-w. PMID: 36959176; PMCID: PMC10036610. ↥
Melchitzky DS, Lewis DA (2000): Tyrosine hydroxylase- and dopamine transporter-immunoreactive axons in the primate cerebellum. Evidence for a lobular- and laminar-specific dopamine innervation. Neuropsychopharmacology. 2000 May;22(5):466-72. doi: 10.1016/S0893-133X(99)00139-6. PMID: 10731621. ↥ ↥
Isaac V, Lopez V, Escobar MJ (2025): Can attention-deficit/hyperactivity disorder be considered a form of cerebellar dysfunction? Front Neurosci. 2025 Jan 22;19:1453025. doi: 10.3389/fnins.2025.1453025. PMID: 39911701; PMCID: PMC11794510. ↥ ↥
Giompres P, Delis F (2005): Dopamine transporters in the cerebellum of mutant mice. Cerebellum. 2005;4(2):105-11. doi: 10.1080/14734220510007851. PMID: 16035192. REVIEW ↥
Bègue I, Elandaloussi Y, Delavari F, Cao H, Moussa-Tooks A, Roser M, Coupé P, Leboyer M, Kaiser S, Houenou J, Brady R, Laidi C (2024): The Cerebellum and Cognitive Function: Anatomical Evidence from a Transdiagnostic Sample. Cerebellum. 2024 Aug;23(4):1399-1410. doi: 10.1007/s12311-023-01645-y. PMID: 38151675; PMCID: PMC11269336. ↥
Joseph B, Nandhu MS, Paulose CS (2010): Dopamine D1 and D2 receptor functional down regulation in the cerebellum of hypoxic neonatal rats: neuroprotective role of glucose and oxygen, epinephrine resuscitation. Pharmacol Res. 2010 Feb;61(2):136-41. doi: 10.1016/j.phrs.2009.08.007. PMID: 19720148. ↥
Locke TM, Fujita H, Hunker A, Johanson SS, Darvas M, du Lac S, Zweifel LS, Carlson ES (2020): Purkinje Cell-Specific Knockout of Tyrosine Hydroxylase Impairs Cognitive Behaviors. Front Cell Neurosci. 2020 Jul 29;14:228. doi: 10.3389/fncel.2020.00228. PMID: 32848620; PMCID: PMC7403473. ↥
Parkkinen S, Radua J, Andrews DS, Murphy D, Dell’Acqua F, Parlatini V (2024): Cerebellar network alterations in adult attention-deficit/hyperactivity disorder. J Psychiatry Neurosci. 2024 Jul 3;49(4):E233-E241. doi: 10.1503/jpn.230146. PMID: 38960626; PMCID: PMC11230668. ↥
Manto M, Adamaszek M, Apps R, Carlson E, Guarque-Chabrera J, Heleven E, Kakei S, Khodakhah K, Kuo SH, Lin CR, Joshua M, Miquel M, Mitoma H, Larry N, Péron JA, Pickford J, Schutter DJLG, Singh MK, Tan T, Tanaka H, Tsai P, Van Overwalle F, Yamashiro K (2024): Consensus Paper: Cerebellum and Reward. Cerebellum. 2024 Oct;23(5):2169-2192. doi: 10.1007/s12311-024-01702-0. PMID: 38769243. REVIEW ↥
Cundari M, Vestberg S, Gustafsson P, Gorcenco S, Rasmussen A (2023): Neurocognitive and cerebellar function in ADHD, autism and spinocerebellar ataxia. Front Syst Neurosci. 2023 Jun 21;17:1168666. doi: 10.3389/fnsys.2023.1168666. Erratum in: Front Syst Neurosci. 2024 Aug 20;18:1462062. doi: 10.3389/fnsys.2024.1462062. PMID: 37415926; PMCID: PMC10321758. REVIEW ↥ ↥
Miquel M, Nicola SM, Gil-Miravet I, Guarque-Chabrera J, Sanchez-Hernandez A (2019): A Working Hypothesis for the Role of the Cerebellum in Impulsivity and Compulsivity. Front Behav Neurosci. 2019 May 7;13:99. doi: 10.3389/fnbeh.2019.00099. PMID: 31133834; PMCID: PMC6513968. ↥
Kim, Woo, Lee, Yoon (2017): Decreased Glial GABA and Tonic Inhibition in Cerebellum of Mouse Model for Attention-Deficit/Hyperactivity Disorder (ADHD). Exp Neurobiol. 2017 Aug;26(4):206-212. doi: 10.5607/en.2017.26.4.206. ↥
Stoodley CJ (2014): Distinct regions of the cerebellum show gray matter decreases in autism, ADHD, and developmental dyslexia. Front Syst Neurosci. 2014 May 20;8:92. doi: 10.3389/fnsys.2014.00092. PMID: 24904314; PMCID: PMC4033133. ↥
Elandaloussi Y, Dufrenne O, Lefebvre A, Houenou J, Senova S, Laidi C (2025): Cerebellar Neuromodulation in Autism Spectrum Disorders and Social Cognition: Insights from Animal and Human Studies. Cerebellum. 2025 Feb 12;24(2):46. doi: 10.1007/s12311-025-01801-6. PMID: 39937336. REVIEW ↥
Xi K, Cai SQ, Yan HF, Tian Y, Cai J, Yang XM, Wang JM, Xing GG (2023): CSMD3 Deficiency Leads to Motor Impairments and Autism-Like Behaviors via Dysfunction of Cerebellar Purkinje Cells in Mice. J Neurosci. 2023 May 24;43(21):3949-3969. doi: 10.1523/JNEUROSCI.1835-22.2023. PMID: 37037606; PMCID: PMC10219040. ↥
Okada NJ, Liu J, Tsang T, Nosco E, McDonald NM, Cummings KK, Jung J, Patterson G, Bookheimer SY, Green SA, Jeste SS, Dapretto M (2022): Atypical cerebellar functional connectivity at 9 months of age predicts delayed socio-communicative profiles in infants at high and low risk for autism. J Child Psychol Psychiatry. 2022 Sep;63(9):1002-1016. doi: 10.1111/jcpp.13555. PMID: 34882790; PMCID: PMC9177892. ↥
Stoodley CJ, D’Mello AM, Ellegood J, Jakkamsetti V, Liu P, Nebel MB, Gibson JM, Kelly E, Meng F, Cano CA, Pascual JM, Mostofsky SH, Lerch JP, Tsai PT (2017): Altered cerebellar connectivity in autism and cerebellar-mediated rescue of autism-related behaviors in mice. Nat Neurosci. 2017 Dec;20(12):1744-1751. doi: 10.1038/s41593-017-0004-1. PMID: 29184200; PMCID: PMC5867894. ↥
Jaber M (2017): Le cervelet comme acteur majeur dans les troubles moteurs des syndromes autistiques [The cerebellum as a major player in motor disturbances related to Autistic Syndrome Disorders]. Encephale. 2017 Apr;43(2):170-175. French. doi: 10.1016/j.encep.2016.03.018. PMID: 27616580. REVIEW ↥
Mosconi MW, Wang Z, Schmitt LM, Tsai P, Sweeney JA (2015): The role of cerebellar circuitry alterations in the pathophysiology of autism spectrum disorders. Front Neurosci. 2015 Sep 1;9:296. doi: 10.3389/fnins.2015.00296. PMID: 26388713; PMCID: PMC4555040. REVIEW ↥ ↥
McKimm E, Corkill B, Goldowitz D, Albritton LM, Homayouni R, Blaha CD, Mittleman G (2014): Glutamate dysfunction associated with developmental cerebellar damage: relevance to autism spectrum disorders. Cerebellum. 2014 Jun;13(3):346-53. doi: 10.1007/s12311-013-0541-4. PMID: 24307139; PMCID: PMC4060592. ↥
Rogers TD, Dickson PE, McKimm E, Heck DH, Goldowitz D, Blaha CD, Mittleman G (2013): Reorganization of circuits underlying cerebellar modulation of prefrontal cortical dopamine in mouse models of autism spectrum disorder. Cerebellum. 2013 Aug;12(4):547-56. doi: 10.1007/s12311-013-0462-2. PMID: 23436049; PMCID: PMC3854915. ↥
Mittleman G, Goldowitz D, Heck DH, Blaha CD (2008): Cerebellar modulation of frontal cortex dopamine efflux in mice: relevance to autism and schizophrenia. Synapse. 2008 Jul;62(7):544-50. doi: 10.1002/syn.20525. PMID: 18435424; PMCID: PMC3854870. ↥ ↥
Wu T, Hallett M (2013): The cerebellum in Parkinson’s disease. Brain. 2013 Mar;136(Pt 3):696-709. doi: 10.1093/brain/aws360. PMID: 23404337; PMCID: PMC7273201. REVIEW ↥
Lewis MM, Galley S, Johnson S, Stevenson J, Huang X, McKeown MJ (2013): The role of the cerebellum in the pathophysiology of Parkinson’s disease. Can J Neurol Sci. 2013 May;40(3):299-306. doi: 10.1017/s0317167100014232. PMID: 23603164; PMCID: PMC6939223. REVIEW ↥
Wu T, Hallett M (2013): Reply: The cerebellum in Parkinson’s disease and parkinsonism in cerebellar disorders. Brain. 2013 Sep;136(Pt 9):e249. doi: 10.1093/brain/awt100. PMID: 23739172. ↥
Simioni AC, Dagher A, Fellows LK (2015): Compensatory striatal-cerebellar connectivity in mild-moderate Parkinson’s disease. Neuroimage Clin. 2015 Nov 11;10:54-62. doi: 10.1016/j.nicl.2015.11.005. PMID: 26702396; PMCID: PMC4669533. ↥
Mirdamadi JL (2016): Cerebellar role in Parkinson’s disease. J Neurophysiol. 2016 Sep 1;116(3):917-9. doi: 10.1152/jn.01132.2015. PMID: 26792889; PMCID: PMC5009206. REVIEW ↥
Parker KL, Narayanan NS, Andreasen NC (2014): The therapeutic potential of the cerebellum in schizophrenia. Front Syst Neurosci. 2014 Sep 15;8:163. doi: 10.3389/fnsys.2014.00163. PMID: 25309350; PMCID: PMC4163988. ↥
Miquel M, Vazquez-Sanroman D, Carbo-Gas M, Gil-Miravet I, Sanchis-Segura C, Carulli D, Manzo J, Coria-Avila GA (2016): Have we been ignoring the elephant in the room? Seven arguments for considering the cerebellum as part of addiction circuitry. Neurosci Biobehav Rev. 2016 Jan;60:1-11. doi: 10.1016/j.neubiorev.2015.11.005. PMID: 26602022. REVIEW ↥
Miquel M, Toledo R, García LI, Coria-Avila GA, Manzo J (2009): Why should we keep the cerebellum in mind when thinking about addiction? Curr Drug Abuse Rev. 2009 Jan;2(1):26-40. doi: 10.2174/1874473710902010026. PMID: 19630735. REVIEW ↥
Vazquez-Sanroman D, Carbo-Gas M, Leto K, Cerezo-Garcia M, Gil-Miravet I, Sanchis-Segura C, Carulli D, Rossi F, Miquel M (2015): Cocaine-induced plasticity in the cerebellum of sensitised mice. Psychopharmacology (Berl). 2015 Dec;232(24):4455-67. doi: 10.1007/s00213-015-4072-1. PMID: 26482898. ↥
Vazquez-Sanroman D, Leto K, Cerezo-Garcia M, Carbo-Gas M, Sanchis-Segura C, Carulli D, Rossi F, Miquel M (2015): The cerebellum on cocaine: plasticity and metaplasticity. Addict Biol. 2015 Sep;20(5):941-55. doi: 10.1111/adb.12223. PMID: 25619460. ↥
Miquel M, Gil-Miravet I, Guarque-Chabrera J (2020): The Cerebellum on Cocaine. Front Syst Neurosci. 2020 Oct 21;14:586574. doi: 10.3389/fnsys.2020.586574. PMID: 33192350; PMCID: PMC7641605. REVIEW ↥
Iversen L, Iversen F, Bloom S, Roth R (2009): Introduction to Neuropsychopharmacology, S. 181 ↥ ↥
Roy S, Field GD (2019): Dopaminergic modulation of retinal processing from starlight to sunlight. J Pharmacol Sci. 2019 May;140(1):86-93. doi: 10.1016/j.jphs.2019.03.006. PMID: 31109761. ↥
Prendergast BJ, Zucker I (2016): Ultradian rhythms in mammalian physiology and behavior. Curr Opin Neurobiol. 2016 Oct;40:150-154. doi: 10.1016/j.conb.2016.07.011. PMID: 27568859. REVIEW ↥
Goh GH, Maloney SK, Mark PJ, Blache D (2019): Episodic Ultradian Events-Ultradian Rhythms. Biology (Basel). 2019 Mar 14;8(1):15. doi: 10.3390/biology8010015. PMID: 30875767; PMCID: PMC6466064. REVIEW ↥
Yao T, Kim R (2025): Mathematical modeling of dopamine rhythms and timing of dopamine reuptake inhibitors. PLoS Comput Biol. 2025 Sep 25;21(9):e1013508. doi: 10.1371/journal.pcbi.1013508. PMID: 40997123; PMCID: PMC12494296. ↥ ↥ ↥ ↥ ↥
Oster H, Damerow S, Kiessling S, Jakubcakova V, Abraham D, Tian J, Hoffmann MW, Eichele G (2006): The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell Metab. 2006 Aug;4(2):163-73. doi: 10.1016/j.cmet.2006.07.002. PMID: 16890544. ↥
Korshunov KS, Blakemore LJ, Trombley PQ (2017): Dopamine: A Modulator of Circadian Rhythms in the Central Nervous System. Front Cell Neurosci. 2017 Apr 3;11:91. doi: 10.3389/fncel.2017.00091. PMID: 28420965; PMCID: PMC5376559. REVIEW ↥
Doyle SE, Grace MS, McIvor W, Menaker M (2002): Circadian rhythms of dopamine in mouse retina: the role of melatonin. Vis Neurosci. 2002 Sep-Oct;19(5):593-601. doi: 10.1017/s0952523802195058. PMID: 12507326. ↥
Chung S, Lee EJ, Yun S, Choe HK, Park SB, Son HJ, Kim KS, Dluzen DE, Lee I, Hwang O, Son GH, Kim K (2014): Impact of circadian nuclear receptor REV-ERBα on midbrain dopamine production and mood regulation. Cell. 2014 May 8;157(4):858-68. doi: 10.1016/j.cell.2014.03.039. PMID: 24813609. ↥
Colita CI, Hermann DM, Filfan M, Colita D, Doepnner TR, Tica O, Glavan D, Popa-Wagner A (2024): Optimizing Chronotherapy in Psychiatric Care: The Impact of Circadian Rhythms on Medication Timing and Efficacy. Clocks Sleep. 2024 Nov 5;6(4):635-655. doi: 10.3390/clockssleep6040043. PMID: 39584972; PMCID: PMC11586979. REVIEW ↥ ↥
Verwey M, Dhir S, Amir S (2016): Circadian influences on dopamine circuits of the brain: regulation of striatal rhythms of clock gene expression and implications for psychopathology and disease. F1000Res. 2016 Aug 24;5:F1000 Faculty Rev-2062. doi: 10.12688/f1000research.9180.1. PMID: 27635233; PMCID: PMC5007753. REVIEW ↥
Wesensten NJ, Belenky G, Kautz MA, Thorne DR, Reichardt RM, Balkin TJ (2002): Maintaining alertness and performance during sleep deprivation: modafinil versus caffeine. Psychopharmacology (Berl). 2002 Jan;159(3):238-47. doi: 10.1007/s002130100916. PMID: 11862356. ↥
McClung CA, Sidiropoulou K, Vitaterna M, Takahashi JS, White FJ, Cooper DC, Nestler EJ (2005): Regulation of dopaminergic transmission and cocaine reward by the Clock gene. Proc Natl Acad Sci U S A. 2005 Jun 28;102(26):9377-81. doi: 10.1073/pnas.0503584102. PMID: 15967985; PMCID: PMC1166621. ↥
McGeer EG, McGeer PL (1966): Circadian rhythm in pineal tyrosine hydroxylase. Science. 1966 Jul 1;153(3731):73-4. doi: 10.1126/science.153.3731.73. PMID: 5938420. ↥
Ferris MJ, España RA, Locke JL, Konstantopoulos JK, Rose JH, Chen R, Jones SR (2014): Dopamine transporters govern diurnal variation in extracellular dopamine tone. Proc Natl Acad Sci U S A. 2014 Jul 1;111(26):E2751-9. doi: 10.1073/pnas.1407935111. PMID: 24979798; PMCID: PMC4084435. ↥
Zawilska JB (1994): The role of dopamine in the regulation of melatonin biosynthesis in vertebrate retina. Acta Neurobiol Exp (Wars). 1994;54 Suppl:47-56. PMID: 7801792. REVIEW ↥
González S, Moreno-Delgado D, Moreno E, Pérez-Capote K, Franco R, Mallol J, Cortés A, Casadó V, Lluís C, Ortiz J, Ferré S, Canela E, McCormick PJ (2012): Circadian-related heteromerization of adrenergic and dopamine D₄ receptors modulates melatonin synthesis and release in the pineal gland. PLoS Biol. 2012;10(6):e1001347. doi: 10.1371/journal.pbio.1001347. PMID: 22723743; PMCID: PMC3378626. ↥
Sellix MT, Freeman ME (2003): Circadian rhythms of neuroendocrine dopaminergic neuronal activity in ovariectomized rats. Neuroendocrinology. 2003 Jan;77(1):59-70. doi: 10.1159/000068334. PMID: 12624542. ↥
Hampp G, Ripperger JA, Houben T, Schmutz I, Blex C, Perreau-Lenz S, Brunk I, Spanagel R, Ahnert-Hilger G, Meijer JH, Albrecht U (2008): Regulation of monoamine oxidase A by circadian-clock components implies clock influence on mood. Curr Biol. 2008 May 6;18(9):678-83. doi: 10.1016/j.cub.2008.04.012. PMID: 18439826. ↥
Taufique SKT, Ehichioya DE, Pendergast JS, Yamazaki S (2022): Genetics and functional significance of the understudied methamphetamine sensitive circadian oscillator (MASCO). F1000Res. 2022 Sep 8;11:1018. doi: 10.12688/f1000research.125432.2. PMID: 36226037; PMCID: PMC9539084. ↥
Abarca C, Albrecht U, Spanagel R (2002): Cocaine sensitization and reward are under the influence of circadian genes and rhythm. Proc Natl Acad Sci U S A. 2002 Jun 25;99(13):9026-30. doi: 10.1073/pnas.142039099. PMID: 12084940; PMCID: PMC124417. ↥