
2. Die 6 dopaminergen Systeme des Gehirns
Das Gehirn beinhaltet etliche Kommunikationssysteme, mittels derer bestimmte Gehirnareale untereinander Informationen austauschen (ähnlich wie Autobahnen innerhalb des gesamten Straßennetzes) und die jeweils bestimmte Neurotransmitter nutzen.
Fünf dieser Kommunikationssysteme basieren auf einem Informationsaustausch mittels Dopamin (dopaminerge Pfade):1
- Mesolimbisches Dopaminsystem (2.2.)
- Mesokortikales Dopaminsystem (2.3.)
- Mesostriatales (nigrostriatales) Dopaminsystem (2.4.)
- Tuberoinfundibuläres Dopaminsystem (2.5.)
- Inzertohypothalamisches Dopaminsystem (2.6.)
Recht jung ist die Erkenntnis, dass das Cerebellum (Kleinhirn) ein eigenes dopaminerges System enthält:
- Cerebellares Dopaminsystem (2.7.)
Daneben besteht noch das dopaminerge System der Retina, über das helles Tageslicht die Dopaminsynthese in der Retina steuert (2.8.1.), was wiederum den circadianen Rhythmus beeinflusst, sowie dopaminerge Zellen im Riechkolben (2.8.2.).
Einige Stimmen bezweifeln die funktionelle Unterscheidung zwischen mesokortikolimbischem und nigrostriatalem Weg,2 z.B. aufgrund von Hinweisen auf eine Steuerung von Belohnung und Aversion durch die Substantia Nigra pars compacta, was darauf hindeutet, dass diese Struktur eine Rolle bei der Belohnung spielt, obwohl sie nicht Teil des mesokortikolimbischen Systems ist, das als Verstärkungssystem verstanden wird.3
Innerhalb der verschiedenen Dopaminsysteme kodieren Dopaminspiegeländerung in einzelnen Gehirnregionen unterschiedliche Verhaltensweisen.
Beispielsweise findet sich bei Ratten, die durch ein Labyrinth laufen, ein stetiger Anstieg des Dopaminspiegels im Striatum, der sein Maximum am Ausgang des Labyrinths hatte.4 Möglicherweise kodiert dies die Zeitabschätzung bis zur erwarteten Belohnung.
Ein hoher Dopaminspiegel in der (beidseitigen) Insula reduziert dagegen die Bereitschaft, Anstrengungen zu unternehmen, um Belohnungen zu erhalten. Der Insula wird die Funktion zugeschrieben, die Kosten einer Anstrengung zu bewerten.5
2.1. Dopaminneuronen im Gehirn
Dopaminneuronen können durch Feststellung ihrer Tyrosinhydroxylase-Immunreaktivität identifiziert werden:6
- Mäuse: 21.000 bis 30.000 TH-positive Neuronen
- Ratten: 45.000 TH-positive Neuronen
- Primaten: 160.000 bis 320.000 TH-immunreaktive Neuronen
- menschliches Mittelhirn: 400.000 bis 600.000 TH-positive Neuronen (Dopaminneuronen)78
- knapp 600.000 bei Vierzigjährigen
- rund 350.000 bei Sechzigjährigen
Die meisten dopaminergen Neurone sind Projektionsneurone, deren lange und weitverzweigte Fortsätzen mehr als hunderttausend Synapsen ausbilden und dadurch zeitgleich ganze Zellgruppen in vielen unterschiedlichen Hirnregionen beeinflussen können. Daher haben sie sehr starken Einfluss auf neuronale Prozesse und Verhalten.7 Humane Dopamin-Neuronen der Substantia nigra sollen über eine Million Synapsen bilden und eine Axon-Gesamtlänge von mehr als 4 m haben können9 und dementsprechend mehrere Jahr benötigen, um auszuwachsen. Dementsprechend bewirken Gehirnentwicklungsstörungen häufig Störungen im dopaminergen System.7
Dopaminerge Nervenzellen finden sich:10
- im ventralen Mittelhirn (überwiegend), daneben auch im ventralen Diencephalon
- im Telencephalon
- im Zwischenhirn
–> Hemmung der Prolaktin-Produktion- hypothalamischer Nucleus arcuatus, A12
- subparafaszikulärer Thalamuskern, A11
–> Innervation des superioren Olivary-Komplex und des inferioren Colliculus im Hirnstamm, A13 (Regulation der auditorische Verarbeitung) - medialer und anertiorer Hypothalamus, A14
- im Mittelhirn
- in der rostralen Hälfte des periaquäduktalen Grau (Substantia grisea periaquaeductalis, zentrales Höhlengrau) des Mittelhirns findet sich bei Primaten eine sehr kleine dopaminerge Zellgruppe, die Aaq genannt wird7
A1512
- existiert nur bei einigen wenigen Spezies (z.B. bei Schafen, Nagetieren, Primaten)
- ist immunreaktiv für Tyrosinhydroxylase, eine Vorstufe von Dopamin
- befindet sich
- in ventralen und dorsalen Komponenten innerhalb des präoptischen periventrikulären Kerns
- in angrenzenden Teilen der anterioren hypothalamischen Region
- A15 schließt kaudal an die dopaminerge Gruppe A14 an.
Die Spezifizierung, Differenzierung und Reifung dopaminerger Neuronen im ventralen Mittelhirn ist ein komplexer Prozess, beeinflusst durch verschiedene Mechanismen, wie:10
- Neurulation
- Proliferation
- Differenzierung von Vorläuferzellen
- Migration
- Bildung von Synapsen
- Bildung neuronaler Schaltkreisen
Diese Mechanismen, die an der Steuerung der dopaminergen Funktionen beteiligt sind, werden durch externe Signale gesteuert, wie
- Morphogene
- Wachstumsfaktoren
- Aktivierung spezifischer Genkaskaden
- Aktivierung zellulärer Interaktionen
2.2. Das mesolimbische Dopaminsystem
Teil des dopaminergen Fokussierungs-, Verstärkungs- und Motivationssystems (zuweilen auch mesencephales dopaminerges System oder mesolimbokortikales System genannt; weiterer Teil: das mesokortikale System).1314
Das mesolimbische, das mesokortikale und das nigrostriatale Dopaminsystem besteht aus dopaminergen Neuronen mit sehr langen Axonen, die weit entfernt liegende Regionen des Gehirns adressieren.15
Das mesolimbische Dopaminsystem umfasst dopaminerge Neuronen im ventralen Tegmentum (VTA) des Mittelhirns, in denen Dopamin gebildet wird, das projiziert wird zum:1617
- Nucleus accumbens im ventralen Striatum
- Hippocampus (Teil des limbischen Systems)
- Amygdala (Teil des limbischen Systems)
- Septum
Das limbische System steuert das emotionale Erleben, dessen Ausdruck (Lust / Unlust) und die Belohnungsverarbeitung.
Dopaminmangel im oder Verletzungen des Nucleus accumbens bewirken eine verringerte Fähigkeit zum Belohnungsaufschub.18
Dopamin steuert im mesolimbischen System motorische Verhaltensabläufe im Kontext von Belohnung (Annäherung an begehrte Dinge) und Reaktion auf neuartige Reize.19
Fehlfunktionen des mesolimbischen Systems:
- bei ADHS:
- Probleme der Verstärkungsmechanismen20
- Belohnungs-Aufschub-Aversion (Abwertung späterer Belohnung)16
- Verzögerungsaversion, Ungeduld20
- Frustrationstoleranz verringert16
- Hyperaktivität, insbesondere in neuen Situationen2021
- Impulsivität2021
- Störungen der Verhaltensinhibition/Verhaltensunterdrückung20
- wechselhaftes Verhalten20
- Störung der Daueraufmerksamkeit20
- bei Schizophrenie aufgrund Dopaminüberschuss:
Aktivierung durch24
- zentrale Stimulanzien
- Nikotin
- Apomorphin
- Amphetamine
- Kokain
- gemischt inhibierend-stimulierende bzw. euphorisierende Substanzen
- Alkohol
- Cannabis
- Opioide
2.3. Das mesokortikale Dopaminsystem
Zweiter Teil des dopaminergen Fokussierungs-, Verstärkungs- und Motivationssystems (zuweilen auch mesencephales dopaminerges System genannt; erster Teil: das mesolimbische System).14
Das mesolimbische, das mesokortikale und das nigrostriatale Dopaminsystem besteht aus dopaminergen Neuronen mit sehr langen Axonen, die weit entfernt liegende Regionen des Gehirns adressieren.15
Es umfasst Verbindungen vom VTA im Brodmann-Areal 10 des Mittelhirnes, in dem Dopamin gebildet wird, zum251416
- PFC
- bei ADHS die wichtigste mesokortikale Gehirnregion
- orbitofrontalen Cortex (OFC)
- ventralen Gyrus cinguli
wo die Dopaminfreisetzung erfolgt.
Fehlfunktionen des mesokortikalen Systems:
- Bei ADHS:
- bei Schizophrenie durch Dopaminmangel hier:
- Aufmerksamkeitsstörungen (Positivsymptom)26
- Affektverflachung (Negativsymptom)26
- Alogie (Denkstörung mit Sprachverarmung, Spracharmut und verlängerter Antwortzeit)26
- Apathie = Teilnahmslosigkeit, mangelnde Erregbarkeit (nicht sexuell)26
- Anhedonie27
Anhedonie (Genussunfähigkeit, verringerte Freudeempfindung) ist auch bei ADHS häufig.
Dopaminmangel im mesokortikalen System führt zu Dopaminüberschuss im nigrostriatalen System, was weitere Hyperaktivität und Impulsprobleme verursacht.28
Aktivierung durch14
- zentrale Stimulanzien
- Nikotin
- Apomorphin
- Amphetamine
- Kokain
- gemischt inhibierend-stimulierende bzw. euphorisierende Substanzen
- Alkohol
- Cannabis
- Opioide
Nur das ADHS-Symptom der mangelnden Inhibition der exekutiven Funktionen wird dopaminerg durch die Basalganglien (Striatum, Putamen) verursacht, während die mangelnde Inhibition der Emotionsregulierung noradrenerg durch den Hippocampus verursacht wird.29 Daher dürfte ersteres einer dopaminergen Behandlung besser zugänglich sein, während Emotionsregulierung und Affektkontrolle besser noradrenerg zu behandeln sein dürften. Dies deckt sich mit unserer Erfahrung, dass Atomoxetin die emotionale Dysregulation bei ADHS besser optimiert als Stimulanzien.
Mesokortikale DA-Neuronen haben im Vergleich zu mesolimbischen oder nigrostriatalen Dopamin-Neuronen
- höhere tonische Feuerrate303132
- mesopräfrontale Dopamin-Neuronen hatten mittlere Entladungsraten von 9,3 Spikes/s und intensive Burst-Aktivität
- mesocinguläre Dopamin-Neuronen 5,9 Spikes/s und intensive Burst-Aktivität
- mesopiriforme Dopamin-Neuronen 4,3 Spikes/s und mäßige Burst-Aktivität
- nigrostriatale Dopamin-Neuronen 3,1 Spikes/s und mäßige Burst-Aktivität
- häufigeres Burst Firing333132
- höhere Dopaminumsatzrate (2 bis 4-fach)343132
- eine stark verringerte Ansprechbarkeit auf DA-Agonisten und -Antagonisten3132
- eine fehlende Toleranz gegenüber der Wirkung von chronisch verabreichten DA-Antagonisten3132
- eine selektive Aktivierung durch Stress35, z.B. Fußschocks31
- verringerte Entwicklung von durch Depolarisation induzierter Inaktivität auf chronische Antipsychotika-Gabe32
- Dopaminsynthese
- D2-Autorezeptoren
2.4. Das nigrostriatale Dopaminsystem
Es umfasst dopaminerge Nervenzellen in der Substantia nigra pars compacta, die in die Basalganglien / in das dorsale Striatum (Nucleus caudatus, Putamen) projizieren,16 und ist hauptsächlich mit der motorischen Kontrolle36 und der Auswahl von Aktionen verbunden.37
Bei ADHS ist das dorsale Striatum die wichtigste nigrostriatale Gehirnregion.
Das mesolimbische, das mesokortikale und das nigrostriatale Dopaminsystem besteht aus dopaminergen Neuronen mit sehr langen Axonen, die weit entfernt liegende Regionen des Gehirns adressieren.15 Humane Dopamin-Neuronen der Substantia nigra könnten mehr als eine Million Synapsen bilden und eine axonale Gesamtlänge von mehr als 4 m haben.9
Die dopaminergen Neurone der Substantia Nigra pars compacta „überfluten“ mit ihrem dichten Netzwerk die Zielregionen im dorsalen Striatum mit Dopamin.7 Im Striatum bilden dopaminerge Fasern alle 4 μm en passant Synapsen. Bei einer Halbwertszeit von etwa 75ms kann Dopamin bis zu 12 μm von seiner Freisetzungsstelle entfernt diffundieren.38
Es wurde eine kaskadenartige Spirale von Projektionen zwischen VTA/Substantia nigra und Nucleus accumbens/Striatum beschrieben. Dopamin-Neuronen im medialen VTA senden an mittelgroße stachelige Projektionsneuronen im ventralen Striatum, welche dann zu lateraleren Teilen des VTA/SNc-Komplexes zurückprojizieren, von wo aus Projektionen zu dorsaleren Teilen des Striatums senden.39
Fehlfunktionen des nigrostriatalen Systems:
- Bei ADHS:
- Bei Chorea Huntington:
- Hyperkinetische Bewegungsstörungen42
- Ticstörungen42
- Bei Parkinson durch Dopaminmangel oder bei Blockade von Dopaminrezeptoren durch Antipsychotika in diesem Bereich:
- Tremor
- Rigor (Muskelstarre, Muskelsteifheit)
- Hypokinese (Bewegungsarmut; Verlangsamung der Bewegungen, eingeschränkte Mimik)
- Akinese
2.5. Das tuberoinfundibuläre System
Das tuberoinfundibuläre Dopaminsystem umfasst Verbindungen vom Nucleus arcuatus und Hypothalamus zum Hypophysenvorderlappen.43
Anders als die Dopaminaufnahme im mPFC, Nucleus accumbens und Nucleus Caudatus / Putamen, die dort mit der Menge der vorhandenen Dopaminrezeptoren korreliert, ist die Dopaminaufnahme des neuroendokrinen tuberoinfundibulären Dopaminsystems geringer und langsamer und entspricht jener der Amygdala.44
Das tuberoinfundibuläre System und das inzertohypothalamische Dopaminsystem haben mittellange Axone.45
2.5.1. Dopamin und Prolaktin
Dopamin hemmt die Ausschüttung von Prolaktin.
- Dopaminmangel, z.B. durch blockierte Dopaminrezeptoren im tuberoinfundibulären System, erhöht folglich die Prolaktin-Ausschüttung aus der Hypophyse, der 2. Stufe der HPA-Achse.
- Prolaktin hat einen zirkadianen Rhythmus
- maximale Spiegel während des non-REM-Schlafs
- großer Einfluss auf den Schlaf. (70 bis 80 % der ADHS-Betroffenen leiden an Schlafstörungen.)
- Prolaktin ist ein Regulator der emotionalen Stressreaktion. Bei akuten und chronischen physischen und psychischen Stresssituationen46 und bei Angst47 ist Prolaktin deutlich erhöht.
- Umgekehrt löst ein hoher Prolaktinwert emotionale Instabilität und Angstwahrnehmung aus.
- Prolaktin wird auch beim Orgasmus ausgeschüttet.
- Prolaktin erhöht das Risiko von Brustkrebs.46
Erhöhte Prolaktinwerte (z.B. aufgrund von Dopaminmangel) bewirken:46
- Depressive Verstimmung / Depressionen
- Antriebsarmut
- allgemeine Müdigkeit
- Erschöpfungszustände
- Konzentrationsstörungen
- Schlafstörungen
- Stimmungsschwankungen
- Angstzustände
- Panikattacken
- Unruhe
- Nervosität
- Reizbarkeit
- Schmerzempfindlichkeit
- soziale Fähigkeiten eingeschränkt
- Novelty Seeking / Sensation Seeking verringert
- Wesensveränderungen
Zusammen mit den Symptomen des Dopaminmangels im mesokortikalen System (Anhedonie = leichte Depression, Antriebsarmut) und des daraus folgenden Dopaminüberschusses im nigrostriatalen System (Hyperaktivität, Impulskontrollstörungen) deckt diese Aufzählung nahezu die gesamten typischen ADHS-Symptome ab.
Dies erklärt mit, wieso die den Dopaminhaushalt regulierenden Stimulanzien die ADHS-Symptome so hervorragend behandeln können.
Sonstige Wirkung von Prolaktin:
Beeinflussung der Homöostase:48
- Regulation der humoralen und zellulären Immunantwort sowie bei Autoimmunerkrankungen (Immunmodulation)
- erhöht den Wassertransport durch die Brustzellmembran, Natriumresorption im Dünndarm.
- Förderung der Gefäßneubildung
Beeinflussung des zentralen Nervensystems:48
- Aktivierung dopaminerger Zellen
- dadurch Selbstregulationskreis
- Stimulation des Appetits
- anxiolytisch (angstlösend)
- Stress reduzierend
- Regulation der Oxytocin produzierenden Nervenzellen
- Stimulation der Myelinisierung im Gehirn
2.6. Das inzertohypothalamische Dopaminsystem
Im inzertohypothalamischen Dopaminsystem finden sich die dopaminergen Neuronen im Hypothalamus in den katecholaminergen Arealen A13 und A14. Diese senden ihre dopaminergen Signale an die Hypothalamuskerne (PVN) und den medialen präoptischen Bereich. Das inzertohypothalamische Dopaminsystem steuert verschiedene Funktionen wie Ernährung, Erektionsfähigkeit und Sexualverhalten.49
Das tuberoinfundibuläre System und das inzertohypothalamische Dopaminsystem haben mittellange Axone.45
Eine systemische Gabe von Dopamin-Agonisten durch Mikroinjektion in den paraventrikulären Nukleus des Hypothalamus (PVN) bewirkt bei männlichen Ratten mittels Dopamin-D2-Rezeptor-Aktivierung eine Peniserektion. Bei Mikroinjektion in den medialen präoptischen Bereich erleichtern sie das Kopulationsverhalten. Es handelt sich um eine Aktivierung des inzertohypothalamischen dopaminergen Systems, dessen Neuronen ihren Ursprung in den katecholaminergen Zellgruppen A13 und A14 des Hypothalamus haben.49
2.7. Das cerebellare Dopaminsystem
Das menschliche Cerebellum beinhaltet 60 bis 80 % aller Neuronen des Gehirns.
Bei ADHS sollen insbesondere involviert sein:5051, wobei die Volumenverringerung bei ADHS genausowenig statistisch signifikant war wie für irgendeine andere Gehirnregion52
- Hinterer inferiorer Kleinhirnwurm (Vermis)
- Hintere inferiore Läppchen
Veränderungen des Kleinhirns können beeinträchtigen:53
- visuell-räumliche Verarbeitung
- sprachliche Fähigkeiten
- Agrammatismus,
- Intonationsveränderungen (Dysprosodie)
- Syntaxprobleme
- verminderter Redefluss
- hintere Regionen des Cerebellums
- Exekutivfunktionen53 54
- Planung
- Aufgabenorganisation
- Handlungsabfolge
- Redefluss
- Arbeitsgedächtnis
- abstraktes Denken
- Problemlösungsstrategien
- Aufgabenwechsel (Taskwechsel)
- gleichzeitige Bewältigung mehrerer Aufgaben
- emotionales Verhalten53 54
- sozial unangemessene Verhaltensweisen
- Stimmungsstörungen
- Inhibitionsprobleme
- zwanghafte Verhaltensweisen
- Exekutivfunktionen53 54
Dopaminerge Mittelgehirnkerne senden ins Cerebellum:55
- aus VTA (A10)
- aus dem Nucleus retrorubralis (A8), schwächer
- aus der Substantia nigra pars compacta (SNpc) (A9), schwächer
Das Cerebellum sendet an diese dopaminerg zurück:55
- aus der vermianen Kleinhirnrinde und aus dem Nucleus fastigialis an das ipsilaterale VTA
- aus dem Nucleus interpositus und dem Nucleus dentatus an das kontralaterale dorsale VTA und die mediale und dorsale Substantia nigra pars compacta
- 20 % dieser Fasern waren bilateral
Das Cerebellum (hier Nucleus dentatus, Nucleus fastigialis und Nucleus interpositus posterior) ist mit VTA, SNpc und Nucleus caudatus dopaminerg verschaltet55, ebenso wie das Neocerebellum mit dem PFC dopaminerg verschaltet ist.56
Zusätzlich zu den dichten Verbindungen zwischen Cerebellum und PFC, den Basalganglien und dem VTA57 existiert im Cerebellum ein unabhängiges dopaminerges Netzwerk.5558596061
Subgruppen des Cerebellums zeigen eine strukturelle Spezialisierung für kognitive Funktionen.62
Im Cerebellum finden sich neben etlichen anderen dopaminergen Mechanismen
- hohe DAT-Spiegel, insbesondere im posterior-inferioren Vermis.5559
- alle Dopaminrezeptoren (DRD1 bis DRD5)55
Sauerstoffmangel im Cerebellum neugeborener Ratten bewirkt eine Verringerung des Dopaminspiegels und eine verringerte Expression von DRD1 und DRD2. Eine Gabe von Glukose, Sauerstoff und Adrenalin behebt diese Mängel teilweise wieder.63
Eine selektive Verringerung dopaminerger Purkinje-Neuronen des Cerebellums korrelierte mit spezifischen Beeinträchtigungen kognitiver Funktionen wie Verhaltensflexibilität, Reaktionshemmung und sozialem Erkennungsgedächtnis.64
Mehrere Studien zeigten, dass das Cerebellum an dopaminergen neurologischen und psychiatrischen Störungen beteiligt ist wie55
- ADHS5065666768546970
- MPH wirkt bei ADHS auch auf die Kleinhirnfunktion, was darauf hindeutet, dass Verhaltensänderungen und sogar motorische Veränderungen bei Kindern mit ADHS, die MPH einnehmen, durch Ergebnisse erklärt werden könnten, die das Kleinhirn betreffen.60
- Autismus715772735474757676777879
- Parkinson8081
- Schizophrenie8579
- Drogenabhängigkeit8687888990
- spinozerebelläre Ataxie67
2.8. Weitere dopaminerge Zellen
2.8.1. Retina (A17)
Dopaminerge Zellen in der amakrinen Zellpopulation der Netzhaut, A17, bilden sehr kurze, lokale Axone und verbinden nur die inneren und die äußeren plexiformen Schichten der Retina.91
Eine hohe Dopaminausschüttung in der Netzhaut stellt das Sehen auf Tageslicht (photopisch, Zapfen-Sehen) ein, während eine niedrige Ausschüttung das Sehen auf Nachtlicht (skotopisch, Stäbchen-Sehen) einstellt.92 Dopamin reduziert über den D1-Rezeptor die Horizontalzell-Kopplung, indem Connexine durch Proteinkinase A phosphoryliert werden und dadurch die Pore geschlossen wird.7
Möglicherweise erklärt dies die erhöhte Lichtempfindlichkeit bei ADHS-Betroffenen, da ADHS mit einem verringertem Dopaminspiegel einhergeht,
Helles Tageslicht steuert die Dopaminsynthese in der Retina und beeinflusst so den circadianen Rhythmus.
2.8.2. Riechkolben (A16)
Die periglomerulären Dopaminzellen des Bulbus olfactorius verbinden Mitralzelldendriten in naheliegenden benachbarten Glomeruli miteinander. Auch hier sind die Axone sehr kurz.91
2.9. Dopaminerge circadiane und ultradiane Rhythmen
Verschiedene Einflüsse steuern die Periodik der Dopaminsynthese und des Dopaminsystems:
- externe Reize
- Belohnung
- Stress
- –> vorübergehende Veränderungen
- endogene zirkadiane Rhythmen
- –> ca, 24-stündiges periodisches Verhalten der Dopaminsynthese, -wiederaufnahme und -freisetzung in einzelnen dopaminergen Neuronen
- ultradiane Rhythmen
Ultradiane Rhythmen haben grundsätzlich wichtige Regelfunktionen:95
- Hormonausschüttung
- Schlafphasen
- Verhaltensaktivierung
- Organisation der Verhaltensaktivität
- Steigerung der Reaktionsfähigkeit auf Umweltreize
In VTA, Substantia nigra und Zwischenhirn, wo Dopamin synthetisiert wird, beeinflusst Dopamin nicht nur die genetischen Mechanismen, die die biologischen Uhren steuern, sondern wird auch von ihnen rhythmisch beeinflusst, was auf eine relevante Rolle dieser Hirnareale bei der Aufrechterhaltung der rhythmischen Funktion hindeutet.96
Die Netzhaut von Säugetieren und von Fischen unterliegt einer täglichen Schwankung der Dopaminspiegel mit einem Höhepunkt am Tag und einer Abnahme nachts, wobei dieser Netzhaut-Dopaminzyklus offenbar durch Melatonin beeinflusst wird.9798
Uhr-Gene spielen bei neuronalen Funktionen, insbesondere bei der Regulierung dopaminerger Bahnen, eine entscheidende Rolle. Die zirkadiane Steuerung des Dopaminsystems wird durch kanonische Uhr-Proteine beeinflusst, die die Dopamin-Expression und -Regulierung direkt beeinflussen. REV-ERBalpha (nukleäre Rezeptor-Unterfamilie 1, Gruppe D, Mitglied 1, NR1D1) ist ein zirkadianer nukleärer Rezeptor, der den TTFL negativ reguliert, indem er auf die Bmal1-mRNA abzielt. Dies beeinflusst die Dopaminproduktion im Mittelhirn durch die Unterdrückung der Tyrosinhydroxylase (Th)-mRNA-Synthese und von Cholecystokinin (CCK).99100
Ob bei Primaten die hypothalamischen Kerne die Hauptregulatoren der zirkadianen Physiologie sind, ist fraglich. Chung et al., 2014, fanden bei Kernen, die typischerweise mit rhythmischen Funktionen assoziiert werden, nur eine geringe bis minimale rhythmische Expression von Transkripten, während sich die höchste Anzahl an zirkadian zyklischen Transkripten im PFC fand, gefolgt vom medialen Globus pallidus, dem Putamen, dem lateralen Globus pallidus und den Mammillarkörpern, die Teil der hypothalamischen Kerne sind. Der SCN zeigte dagegen eine sehr geringe Anzahl an zyklischen Transkripten. Zugleich wies diese Studie eine zyklische Expression von Dopamin-D2-Rezeptoren im Putamen nach. Intrinsische autonome zirkadiane Uhren fanden sich in den genannten Hirnarealen mit hoher zyklischer Genaktivität jedoch nicht.100
Die Expression der Uhr-Gene unterliegt Rhythmen, die je nach Gehirnbereich durch unterschiedliche Reize beeinflusst werden:101
- im dorsalen Striatum durch Veränderungen der Dopaminfreisetzung
- dies soll potenzielle Auswirkungen auf die Parkinson-Krankheit und Drogenabhängigkeit haben
- im ventralen Striatum durch psychische und physische Stressoren
- die könne Auswirkungen auf schwere Depressionen haben
- im limbischen Vorderhirn durch psychische und physische Stressoren
- die könne Auswirkungen auf schwere Depressionen haben
Es gibt Hinweise auf tageszeitliche Effekte von Dopamin-Wiederaufnahmehemmern, wonach eine Einnahme zum unpassenden Zeitpunkt starke Dopaminspitzen und anschließende Dopaminabfälle verursachen kann, während eine passende Einnahme den Dopaminspiegel deutlich länger stabil hält. Die Aktivität dopaminerger Neuronen zeigt intrinsische vierstündige ultradiane Rhythmen, die unabhängig von zirkadianen Einflüssen sind.95
Eine Gabe von Dopaminwiederaufnahmehemmern bewirkte in einer mathematischen Modellierung
- während der circadianen Tiefpunkte einen anhaltenden Anstieg der Dopaminspiegel
- die Erhöhung war zudem gleichmäßiger, was für Medikationszwecke vorteilhaft ist
- während der circadianen Spitzen große Schwankungen der Dopaminspiegel
- die Gesamterhöhung des extrazellulären Dopamins war gleich groß, aber sehr viel unregelmäßiger, was für Medikationszwecke nachteilig ist
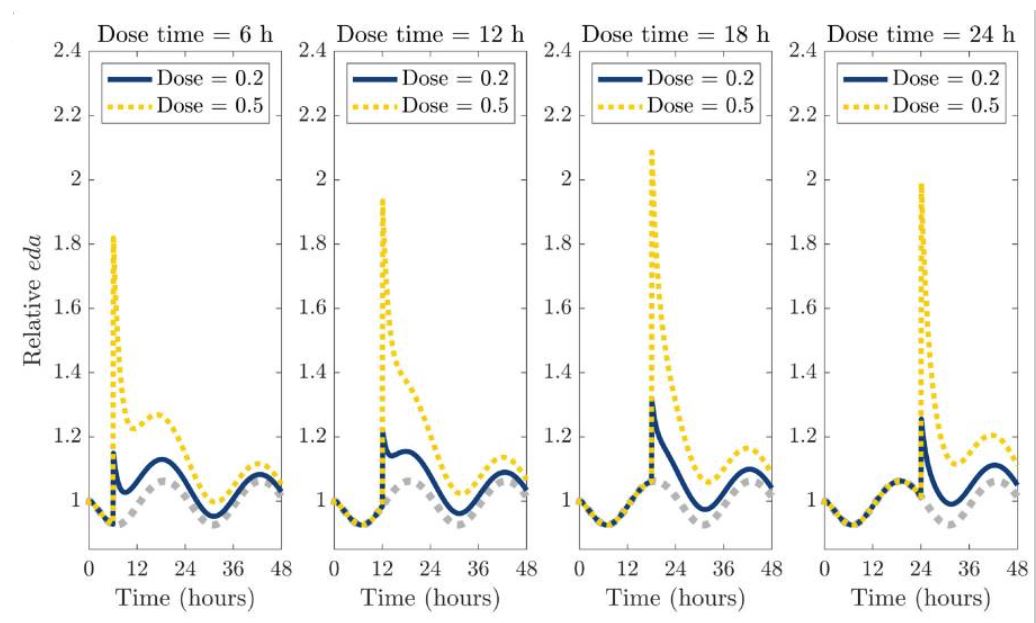
Quelle: Die Grafik mit dem Originaltitel „Single-dose administration of a dopamine reuptake inhibitor (DRI) at different times of the day.“ aus und von Yao, T., & Kim, R. (2025). Mathematical modeling of dopamine rhythms and timing of dopamine reuptake inhibitors. PLoS computational biology, 21(9), e1013508. https://doi.org/10.1371/journal.pcbi.1013508 ist lizenziert unter CC BY 4.0.
Der Tageszeitpunkt der Gabe von Dopaminwiederaufnahmehemmern bestimmte damit auch die Wirkdauer.
- Eine Modafinilgabe während des zirkadianen Dopamin-Tiefpunkts am frühen Morgen, also vor dem natürlichen Anstieg der Dopaminsynthese, zeigte die deutlichste Wirkung auf Wachheit und Leistung bei kognitiven Aufgaben bei Schlafentzug, die zudem 10 Stunden anhielt, obwohl die Halbwertszeit von Modafinil 10 bis 15 Stunden beträgt.102
- der Anstieg des extrazellulären Dopamins blieb relativ gering, aber der zirkadiane Rhythmus setzt ein und der extrazelluläre Dopaminspiegel bleibt länger erhöht95
- Bei einer einmaligen Dopaminwiederaufnahmehemmer-Gabe bei einem erhöhten zirkadianen Dopamin-Wert und bevor die Dopaminsynthese aufgrund des zirkadianen Rhythmus natürlicherweise abnimmt, führt zu einem starken, aber kurzen Anstieg des extrazellulären Dopamins (z. B. 3 bis 6 Stunden)95
- die von dem mathematischen Modell vorhergesagten tageszeitlichen Effekte decken sich mit Ergebnissen klinischer Studien zur kognitiven Leistungsfähigkeit
Der zirkadiane Rhythmus beeinflusst Dopaminsynthese, Dopaminfreisetzung und Dopaminabbau:
- Dopaminsynthese
- Dopaminfreisetzung
- unterliegt einem zirkadianen Rhythmus, was Verhalten und physiologische Rhythmen beeinflusst
- jedoch sollen Dopaminneuronen im Mittelhirn keine tageszeitlichen Schwankungen der Feuerrate aufweisen105
- beeinflusst Melatonin106
- Melatoninfreisetzung aus der Zirbeldrüse
- ist stark rhythmisch
- beruht auf der Heteromerisierung adrenerger Rezeptoren mit Dopamin-D4-Rezeptoren107
- Melatoninfreisetzung aus der Zirbeldrüse
- unterliegt einem zirkadianen Rhythmus, was Verhalten und physiologische Rhythmen beeinflusst
- Dopaminspiegelschwankungen
- auch in hypothalamischen neuroendokrinen Neuronen, die die präzise zeitlich abgestimmte Freisetzung von Fortpflanzungshormonen wie Prolaktin steuern, indem Dopamin Prolaktin hemmt
- Dopamin aus tuberoinfundibulären Neuronen unterliegt einem Aktivitätsrhythmus im Hell-Dunkel-Zyklus, der in konstanter Dunkelheit frei läuft und durch Licht synchronisiert wird, was auf eine zirkadiane Aktivität der tuberoinfundibulären Dopaminneuronenaktivität hindeutet108
- Dopaminabbau
- MAOA, das Dopamin abbaut, wird durch die innere Uhr gesteuert und beeinflusst stimmungsbezogene Verhaltensweisen109
Durch eine (nicht-medikamentöse) Gabe von Stimulanzien kann bei Nagetieren ein zirkadianer Rhythmus hervorgerufen werden, der sogenannte methamphetaminempfindliche zirkadiane Oszillator (MASCO). Dieser hängt offenbar (auch) mit dem dopaminerg gesteuerten Dopamin-Ultradianen Oszillator, DUO, zusammen.110 Es wäre denkbar, dass die zirkadianen Schlafprobleme bei ADHS durch dopaminerge Zusammenhänge mit dem MASCO verbunden sind.
Wiederholten Kokaininjektionen zeigen bei Wildtyp-Mäuse eine sensibilisierte Verhaltensreaktion. Während diese Sensibilisierung bei mPer1-Knockout-Mäusen fehlte, zeigten mPer2-Mutanten eine hypersensibilisierte Reaktion auf Kokain. Experimente zur konditionierten Ortspräferenz ergaben ähnliche Verhaltensreaktionen: mPer1-Knockout-Mäuse zeigten keinerlei Kokainbelohnung, mPer2-Mutanten zeigten eine starke kokaininduzierte Ortspräferenz. Bei C57/BL6J-Mäusen zeigte sich bei Kokaingaben zu unterschiedlichen Uhrzeiten, dass die durch Kokain induzierte Verhaltenssensibilisierung und die Ortspräferenz unter der Kontrolle der circadianen Uhr stehen.111
Rensing, Koch, Rippe, Rippe (2006): Der Mensch im Stress; Psyche, Körper, Moleküle, Kapitel 4: neurobiologische Grundlagen von Stressreaktionen, Seite 89 ↥
Koevoet D, Deschamps PKH, Kenemans JL (2023): Catecholaminergic and cholinergic neuromodulation in autism spectrum disorder: A comparison to attention-deficit hyperactivity disorder. Front Neurosci. 2023 Jan 6;16:1078586. doi: 10.3389/fnins.2022.1078586. PMID: 36685234; PMCID: PMC9853424. REVIEW ↥
Ilango A, Kesner AJ, Keller KL, Stuber GD, Bonci A, Ikemoto S (2014): Similar roles of substantia nigra and ventral tegmental dopamine neurons in reward and aversion. J Neurosci. 2014 Jan 15;34(3):817-22. doi: 10.1523/JNEUROSCI.1703-13.2014. PMID: 24431440; PMCID: PMC3891961. ↥
Howe, Tierney, Sandberg, Phillips, Graybiel (2013): Prolonged Dopamine Signalling in Striatum Signals Proximity and Value of Distant Rewards; Nature. 2013 Aug 29; 500(7464): 575–579. doi: 10.1038/nature12475; PMCID: PMC3927840; NIHMSID: NIHMS507218 ↥
Treadway, Buckholtz, Cowan, Woodward, Li, Ansari, Baldwin, Schwartzman, Kessler, Zald (2012): Dopaminergic Mechanisms of Individual Differences in Human Effort-Based Decision-Making; Journal of Neuroscience 2 May 2012, 32 (18) 6170-6176; DOI: http://dx.doi.org/10.1523/JNEUROSCI.6459-11.2012 ↥
Baik (2020): Stress and the dopaminergic reward system. Exp Mol Med. 2020 Dec;52(12):1879-1890. doi: 10.1038/s12276-020-00532-4. PMID: 33257725; PMCID: PMC8080624. REVIEW ↥
Rillich (2019): Das dopaminerge System im Gehirn des Menschen: molekulare Grundlagen, Anatomie, Physiologie und Pathologie ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥
Azizi SA (2022): Monoamines: Dopamine, Norepinephrine, and Serotonin, Beyond Modulation, “Switches” That Alter the State of Target Networks. Neuroscientist. 2022 Apr;28(2):121-143. doi: 10.1177/1073858420974336. PMID: 33292070. REVIEW ↥ ↥
Pissadaki EK, Bolam JP (2013): The energy cost of action potential propagation in dopamine neurons: clues to susceptibility in Parkinson’s disease. Front Comput Neurosci. 2013 Mar 18;7:13. doi: 10.3389/fncom.2013.00013. PMID: 23515615; PMCID: PMC3600574. ↥ ↥
Speranza L, di Porzio U, Viggiano D, de Donato A, Volpicelli F (2021): Dopamine: The Neuromodulator of Long-Term Synaptic Plasticity, Reward and Movement Control. Cells. 2021 Mar 26;10(4):735. doi: 10.3390/cells10040735. PMID: 33810328; PMCID: PMC8066851. REVIEW ↥ ↥
Pignatelli A, Belluzzi O (2017): Dopaminergic Neurones in the Main Olfactory Bulb: An Overview from an Electrophysiological Perspective. Front Neuroanat. 2017 Feb 14;11:7. doi: 10.3389/fnana.2017.00007. PMID: 28261065; PMCID: PMC5306133. REVIEW ↥
van Vulpen EH, Yang CR, Nissen R, Renaud LP (1999): Hypothalamic A14 and A15 catecholamine cells provide the dopaminergic innervation to the supraoptic nucleus in rat: a combined retrograde tracer and immunohistochemical study. Neuroscience. 1999;93(2):675-80. doi: 10.1016/s0306-4522(99)00173-6. PMID: 10465451. ↥
Genro JP, Kieling C, Rohde LA, Hutz MH (2010): Attention-deficit/hyperactivity disorder and the dopaminergic hypotheses. Expert Rev Neurother. 2010 Apr;10(4):587-601. doi: 10.1586/ern.10.17. PMID: 20367210. REVIEW ↥
Edel, Vollmoeller (2006): ADHS bei Erwachsenen, Seite 110 ↥ ↥ ↥ ↥
Iversen L, Iversen F, Bloom S, Roth R (2009): Introduction to Neuropsychopharmacology, S. 211 ↥ ↥ ↥
Müller, Candrian, Kropotov (2011): ADHS – Neurodiagnostik in der Praxis, Seite 83 ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥
Gatzke-Kopp, Beauchaine (2007): Central nervous system substrates of impulsivity: Implications for the development of attention-deficit/hyperactivity disorder and conduct disorder. In: Coch, Dawson, Fischer ( Eds): Human behavior, learning, and the developing brain: Atypical development. New York: Guilford Press; 2007. pp. 239–263; 246 ↥
Cardinal, Pennicott, Sugathapala, Robbins, Everitt (2001): Impulsive choice induced in rats by lesions of the nucleus accumbens core. Science; (10.1126/science.1060818). ↥
Rensing, Koch, Rippe, Rippe (2006): Der Mensch im Stress; Psyche, Körper, Moleküle, Kapitel 4: neurobiologische Grundlagen von Stressreaktionen, Seite 90 ↥
Sagvolden, Johansen, Aase, Russell (2005): A dynamic developmental theory of attention-deficit/hyperactivity disorder (ADHD) predominantly hyperactive/impulsive and combined subtypes. Behav Brain Sci. 2005 Jun;28(3):397-419; discussion 419-68. ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥
Gatzke-Kopp, Beauchaine (2007): Central nervous system substrates of impulsivity: Implications for the development of attention-deficit/hyperactivity disorder and conduct disorder. In: Coch, Dawson, Fischer ( Eds): Human behavior, learning, and the developing brain: Atypical development. New York: Guilford Press; 2007. pp. 239–263; 253 ↥ ↥ ↥ ↥
Stahl (2000): Essential Psychopharmacology, Neuroscientific Basis and Practical Applications. Second Edition, Cambridge University Press; zitiert nach Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 66 ↥
Stahl (2000): Essential Psychopharmacology, Neuroscientific Basis and Practical Applications. Second Edition, Cambridge University Press, zitiert nach Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 66 ↥
Edel, Vollmoeller (2006): ADHS bei Erwachsenen, Seite 110, mwN ↥
Le Moal, Simon (1991): Mesocorticolimbic dopaminergic network: functional and regulatory roles; Physiological Reviews. 1 January 1991 Vol. 71 no. 1, 155-234 DOI: ↥
Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 66 ↥ ↥ ↥ ↥
Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 66 ↥
Castellanos (1997): Toward a pathophysiology of attention-deficit hyperactivity disorder. Clin. Pediatr. 36, 381-393 ↥
Müller, Candrian, Kropotov (2011): ADHS – Neurodiagnostik in der Praxis, Seite 85 ↥
Chiodo LA, Bannon MJ, Grace AA, Roth RH, Bunney BS (1984): Evidence for the absence of impulse-regulating somatodendritic and synthesis-modulating nerve terminal autoreceptors on subpopulations of mesocortical dopamine neurons. Neuroscience. 1984 May;12(1):1-16. doi: 10.1016/0306-4522(84)90133-7. PMID: 6462443. ↥
Bannon MJ, Roth RH (1983): Pharmacology of mesocortical dopamine neurons. Pharmacol Rev. 1983 Mar;35(1):53-68. PMID: 6138783. REVIEW ↥ ↥ ↥ ↥ ↥ ↥
Iversen L, Iversen F, Bloom S, Roth R (2009): Introduction to Neuropsychopharmacology, S. 208 ↥ ↥ ↥ ↥ ↥ ↥ ↥ ↥
White FJ, Wang RY (1983): Comparison of the effects of chronic haloperidol treatment on A9 and A10 dopamine neurons in the rat. Life Sci. 1983 Feb 28;32(9):983-93. doi: 10.1016/0024-3205(83)90929-3. PMID: 6827927. ↥
Bannon MJ, Bunney EB, Roth RH (1981): Mesocortical dopamine neurons: rapid transmitter turnover compared to other brain catecholamine systems. Brain Res. 1981 Aug 10;218(1-2):376-82. PMID: 7272743. ↥
Finlay, Zigmond (1997): The effects of stress on central dopaminergic neurons: possible clinical implications. Neurochem Res. 1997 Nov;22(11):1387-94. REVIEW ↥ ↥ ↥
Liu, Kaeser (2019): Mechanisms and regulation of dopamine release. Curr Opin Neurobiol. 2019 Aug;57:46-53. doi: 10.1016/j.conb.2019.01.001. PMID: 30769276; PMCID: PMC6629510. ↥
Liu, Goel, Kaeser (2021): Spatial and temporal scales of dopamine transmission. Nat Rev Neurosci. 2021 Jun;22(6):345-358. doi: 10.1038/s41583-021-00455-7. PMID: 33837376; PMCID: PMC8220193. ↥
Seamans JK, Yang CR (2004): The principal features and mechanisms of dopamine modulation in the prefrontal cortex. Prog Neurobiol. 2004 Sep;74(1):1-58. doi: 10.1016/j.pneurobio.2004.05.006. Erratum in: Prog Neurobiol. 2004 Dec;74(5):321. PMID: 15381316. ↥
Morikawa H, Paladini CA (2011): Dynamic regulation of midbrain dopamine neuron activity: intrinsic, synaptic, and plasticity mechanisms. Neuroscience. 2011 Dec 15;198:95-111. doi: 10.1016/j.neuroscience.2011.08.023. PMID: 21872647; PMCID: PMC3221882. REVIEW ↥
Castellanos 1997, zitiert nach Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 67 ↥ ↥
Gatzke-Kopp, Beauchaine (2007): Central nervous system substrates of impulsivity: Implications for the development of attention-deficit/hyperactivity disorder and conduct disorder. In: Coch, Dawson, Fischer (Eds): Human behavior, learning, and the developing brain: Atypical development. New York: Guilford Press; 2007. pp. 239–263; 253 ↥ ↥
Franck (2003): Hyperaktivität und Schizophrenie – eine explorative Studie; Dissertation, Seite 67 ↥ ↥
Jayanti S, Dalla Verde C, Tiribelli C, Gazzin S (2023): Inflammation, Dopaminergic Brain and Bilirubin. Int J Mol Sci. 2023 Jul 14;24(14):11478. doi: 10.3390/ijms241411478. PMID: 37511235; PMCID: PMC10380707. ↥
Garris PA, Wightman RM (1994): Different kinetics govern dopaminergic transmission in the amygdala, prefrontal cortex, and striatum: an in vivo voltammetric study. J Neurosci. 1994 Jan;14(1):442-50. doi: 10.1523/JNEUROSCI.14-01-00442.1994. PMID: 8283249; PMCID: PMC6576851. ↥
Iversen L, Iversen F, Bloom S, Roth R (2009): Introduction to Neuropsychopharmacology, S. 212 ↥ ↥
Prolaktin: Wikipedia, Stand 04.07.2016 ↥ ↥ ↥
Athanasoulia (2011): Prolaktinome und Psyche, Max Planck Institut für Psychatrie ↥
Melis, Sanna, Argiolas (2022): Dopamine, Erectile Function and Male Sexual Behavior from the Past to the Present: A Review. Brain Sci. 2022 Jun 24;12(7):826. doi: 10.3390/brainsci12070826. PMID: 35884633; PMCID: PMC9312911. REVIEW ↥ ↥
Braga-Neto P, Barbosa Dos Santos MW, Scott SSO, Munhoz RP, Novis LE, Pedroso JL, Barsottini OGP (2025): The cerebellum and psychiatric disorders: unraveling its role in mental health. Arq Neuropsiquiatr. 2025 Aug;83(8):1-8. doi: 10.1055/s-0045-1810408. PMID: 40886707. REVIEW ↥ ↥
Cortese S (2012): The neurobiology and genetics of Attention-Deficit/Hyperactivity Disorder (ADHD): what every clinician should know. Eur J Paediatr Neurol. 2012 Sep;16(5):422-33. doi: 10.1016/j.ejpn.2012.01.009. PMID: 22306277. REVIEW ↥
Valera EM, Faraone SV, Murray KE, Seidman LJ (2007): Meta-analysis of structural imaging findings in attention-deficit/hyperactivity disorder. Biol Psychiatry. 2007 Jun 15;61(12):1361-9. doi: 10.1016/j.biopsych.2006.06.011. PMID: 16950217. METASTUDY ↥
Jacobi H, Faber J, Timmann D, Klockgether T (2021): Update cerebellum and cognition. J Neurol. 2021 Oct;268(10):3921-3925. doi: 10.1007/s00415-021-10486-w. PMID: 33656586; PMCID: PMC8463403. ↥ ↥ ↥
Bruchhage MMK, Bucci MP, Becker EBE (2018): Cerebellar involvement in autism and ADHD. Handb Clin Neurol. 2018;155:61-72. doi: 10.1016/B978-0-444-64189-2.00004-4. PMID: 29891077. REVIEW ↥ ↥ ↥ ↥
Flace P, Livrea P, Basile GA, Galletta D, Bizzoca A, Gennarini G, Bertino S, Branca JJV, Gulisano M, Bianconi S, Bramanti A, Anastasi G (2021): The Cerebellar Dopaminergic System. Front Syst Neurosci. 2021 Aug 5;15:650614. doi: 10.3389/fnsys.2021.650614. PMID: 34421548; PMCID: PMC8375553. REVIEW ↥ ↥ ↥ ↥ ↥ ↥ ↥
Habas C (2021): Functional Connectivity of the Cognitive Cerebellum. Front Syst Neurosci. 2021 Apr 8;15:642225. doi: 10.3389/fnsys.2021.642225. PMID: 33897382; PMCID: PMC8060696. REVIEW ↥
Zhou P, Peng S, Wen S, Lan Q, Zhuang Y, Li X, Shi M, Zhang C (2024): The Cerebellum-Ventral Tegmental Area Microcircuit and Its Implications for Autism Spectrum Disorder: A Narrative Review. Neuropsychiatr Dis Treat. 2024 Oct 29;20:2039-2048. doi: 10.2147/NDT.S485487. PMID: 39494383; PMCID: PMC11531233. REVIEW ↥ ↥
Li C, Saliba NB, Martin H, Losurdo NA, Kolahdouzan K, Siddiqui R, Medeiros D, Li W (2023): Purkinje cell dopaminergic inputs to astrocytes regulate cerebellar-dependent behavior. Nat Commun. 2023 Mar 23;14(1):1613. doi: 10.1038/s41467-023-37319-w. PMID: 36959176; PMCID: PMC10036610. ↥
Melchitzky DS, Lewis DA (2000): Tyrosine hydroxylase- and dopamine transporter-immunoreactive axons in the primate cerebellum. Evidence for a lobular- and laminar-specific dopamine innervation. Neuropsychopharmacology. 2000 May;22(5):466-72. doi: 10.1016/S0893-133X(99)00139-6. PMID: 10731621. ↥ ↥
Isaac V, Lopez V, Escobar MJ (2025): Can attention-deficit/hyperactivity disorder be considered a form of cerebellar dysfunction? Front Neurosci. 2025 Jan 22;19:1453025. doi: 10.3389/fnins.2025.1453025. PMID: 39911701; PMCID: PMC11794510. ↥ ↥
Giompres P, Delis F (2005): Dopamine transporters in the cerebellum of mutant mice. Cerebellum. 2005;4(2):105-11. doi: 10.1080/14734220510007851. PMID: 16035192. REVIEW ↥
Bègue I, Elandaloussi Y, Delavari F, Cao H, Moussa-Tooks A, Roser M, Coupé P, Leboyer M, Kaiser S, Houenou J, Brady R, Laidi C (2024): The Cerebellum and Cognitive Function: Anatomical Evidence from a Transdiagnostic Sample. Cerebellum. 2024 Aug;23(4):1399-1410. doi: 10.1007/s12311-023-01645-y. PMID: 38151675; PMCID: PMC11269336. ↥
Joseph B, Nandhu MS, Paulose CS (2010): Dopamine D1 and D2 receptor functional down regulation in the cerebellum of hypoxic neonatal rats: neuroprotective role of glucose and oxygen, epinephrine resuscitation. Pharmacol Res. 2010 Feb;61(2):136-41. doi: 10.1016/j.phrs.2009.08.007. PMID: 19720148. ↥
Locke TM, Fujita H, Hunker A, Johanson SS, Darvas M, du Lac S, Zweifel LS, Carlson ES (2020): Purkinje Cell-Specific Knockout of Tyrosine Hydroxylase Impairs Cognitive Behaviors. Front Cell Neurosci. 2020 Jul 29;14:228. doi: 10.3389/fncel.2020.00228. PMID: 32848620; PMCID: PMC7403473. ↥
Parkkinen S, Radua J, Andrews DS, Murphy D, Dell’Acqua F, Parlatini V (2024): Cerebellar network alterations in adult attention-deficit/hyperactivity disorder. J Psychiatry Neurosci. 2024 Jul 3;49(4):E233-E241. doi: 10.1503/jpn.230146. PMID: 38960626; PMCID: PMC11230668. ↥
Manto M, Adamaszek M, Apps R, Carlson E, Guarque-Chabrera J, Heleven E, Kakei S, Khodakhah K, Kuo SH, Lin CR, Joshua M, Miquel M, Mitoma H, Larry N, Péron JA, Pickford J, Schutter DJLG, Singh MK, Tan T, Tanaka H, Tsai P, Van Overwalle F, Yamashiro K (2024): Consensus Paper: Cerebellum and Reward. Cerebellum. 2024 Oct;23(5):2169-2192. doi: 10.1007/s12311-024-01702-0. PMID: 38769243. REVIEW ↥
Cundari M, Vestberg S, Gustafsson P, Gorcenco S, Rasmussen A (2023): Neurocognitive and cerebellar function in ADHD, autism and spinocerebellar ataxia. Front Syst Neurosci. 2023 Jun 21;17:1168666. doi: 10.3389/fnsys.2023.1168666. Erratum in: Front Syst Neurosci. 2024 Aug 20;18:1462062. doi: 10.3389/fnsys.2024.1462062. PMID: 37415926; PMCID: PMC10321758. REVIEW ↥ ↥
Miquel M, Nicola SM, Gil-Miravet I, Guarque-Chabrera J, Sanchez-Hernandez A (2019): A Working Hypothesis for the Role of the Cerebellum in Impulsivity and Compulsivity. Front Behav Neurosci. 2019 May 7;13:99. doi: 10.3389/fnbeh.2019.00099. PMID: 31133834; PMCID: PMC6513968. ↥
Kim, Woo, Lee, Yoon (2017): Decreased Glial GABA and Tonic Inhibition in Cerebellum of Mouse Model for Attention-Deficit/Hyperactivity Disorder (ADHD). Exp Neurobiol. 2017 Aug;26(4):206-212. doi: 10.5607/en.2017.26.4.206. ↥
Stoodley CJ (2014): Distinct regions of the cerebellum show gray matter decreases in autism, ADHD, and developmental dyslexia. Front Syst Neurosci. 2014 May 20;8:92. doi: 10.3389/fnsys.2014.00092. PMID: 24904314; PMCID: PMC4033133. ↥
Elandaloussi Y, Dufrenne O, Lefebvre A, Houenou J, Senova S, Laidi C (2025): Cerebellar Neuromodulation in Autism Spectrum Disorders and Social Cognition: Insights from Animal and Human Studies. Cerebellum. 2025 Feb 12;24(2):46. doi: 10.1007/s12311-025-01801-6. PMID: 39937336. REVIEW ↥
Xi K, Cai SQ, Yan HF, Tian Y, Cai J, Yang XM, Wang JM, Xing GG (2023): CSMD3 Deficiency Leads to Motor Impairments and Autism-Like Behaviors via Dysfunction of Cerebellar Purkinje Cells in Mice. J Neurosci. 2023 May 24;43(21):3949-3969. doi: 10.1523/JNEUROSCI.1835-22.2023. PMID: 37037606; PMCID: PMC10219040. ↥
Okada NJ, Liu J, Tsang T, Nosco E, McDonald NM, Cummings KK, Jung J, Patterson G, Bookheimer SY, Green SA, Jeste SS, Dapretto M (2022): Atypical cerebellar functional connectivity at 9 months of age predicts delayed socio-communicative profiles in infants at high and low risk for autism. J Child Psychol Psychiatry. 2022 Sep;63(9):1002-1016. doi: 10.1111/jcpp.13555. PMID: 34882790; PMCID: PMC9177892. ↥
Stoodley CJ, D’Mello AM, Ellegood J, Jakkamsetti V, Liu P, Nebel MB, Gibson JM, Kelly E, Meng F, Cano CA, Pascual JM, Mostofsky SH, Lerch JP, Tsai PT (2017): Altered cerebellar connectivity in autism and cerebellar-mediated rescue of autism-related behaviors in mice. Nat Neurosci. 2017 Dec;20(12):1744-1751. doi: 10.1038/s41593-017-0004-1. PMID: 29184200; PMCID: PMC5867894. ↥
Jaber M (2017): Le cervelet comme acteur majeur dans les troubles moteurs des syndromes autistiques [The cerebellum as a major player in motor disturbances related to Autistic Syndrome Disorders]. Encephale. 2017 Apr;43(2):170-175. French. doi: 10.1016/j.encep.2016.03.018. PMID: 27616580. REVIEW ↥
Mosconi MW, Wang Z, Schmitt LM, Tsai P, Sweeney JA (2015): The role of cerebellar circuitry alterations in the pathophysiology of autism spectrum disorders. Front Neurosci. 2015 Sep 1;9:296. doi: 10.3389/fnins.2015.00296. PMID: 26388713; PMCID: PMC4555040. REVIEW ↥ ↥
McKimm E, Corkill B, Goldowitz D, Albritton LM, Homayouni R, Blaha CD, Mittleman G (2014): Glutamate dysfunction associated with developmental cerebellar damage: relevance to autism spectrum disorders. Cerebellum. 2014 Jun;13(3):346-53. doi: 10.1007/s12311-013-0541-4. PMID: 24307139; PMCID: PMC4060592. ↥
Rogers TD, Dickson PE, McKimm E, Heck DH, Goldowitz D, Blaha CD, Mittleman G (2013): Reorganization of circuits underlying cerebellar modulation of prefrontal cortical dopamine in mouse models of autism spectrum disorder. Cerebellum. 2013 Aug;12(4):547-56. doi: 10.1007/s12311-013-0462-2. PMID: 23436049; PMCID: PMC3854915. ↥
Mittleman G, Goldowitz D, Heck DH, Blaha CD (2008): Cerebellar modulation of frontal cortex dopamine efflux in mice: relevance to autism and schizophrenia. Synapse. 2008 Jul;62(7):544-50. doi: 10.1002/syn.20525. PMID: 18435424; PMCID: PMC3854870. ↥ ↥
Wu T, Hallett M (2013): The cerebellum in Parkinson’s disease. Brain. 2013 Mar;136(Pt 3):696-709. doi: 10.1093/brain/aws360. PMID: 23404337; PMCID: PMC7273201. REVIEW ↥
Lewis MM, Galley S, Johnson S, Stevenson J, Huang X, McKeown MJ (2013): The role of the cerebellum in the pathophysiology of Parkinson’s disease. Can J Neurol Sci. 2013 May;40(3):299-306. doi: 10.1017/s0317167100014232. PMID: 23603164; PMCID: PMC6939223. REVIEW ↥
Wu T, Hallett M (2013): Reply: The cerebellum in Parkinson’s disease and parkinsonism in cerebellar disorders. Brain. 2013 Sep;136(Pt 9):e249. doi: 10.1093/brain/awt100. PMID: 23739172. ↥
Simioni AC, Dagher A, Fellows LK (2015): Compensatory striatal-cerebellar connectivity in mild-moderate Parkinson’s disease. Neuroimage Clin. 2015 Nov 11;10:54-62. doi: 10.1016/j.nicl.2015.11.005. PMID: 26702396; PMCID: PMC4669533. ↥
Mirdamadi JL (2016): Cerebellar role in Parkinson’s disease. J Neurophysiol. 2016 Sep 1;116(3):917-9. doi: 10.1152/jn.01132.2015. PMID: 26792889; PMCID: PMC5009206. REVIEW ↥
Parker KL, Narayanan NS, Andreasen NC (2014): The therapeutic potential of the cerebellum in schizophrenia. Front Syst Neurosci. 2014 Sep 15;8:163. doi: 10.3389/fnsys.2014.00163. PMID: 25309350; PMCID: PMC4163988. ↥
Miquel M, Vazquez-Sanroman D, Carbo-Gas M, Gil-Miravet I, Sanchis-Segura C, Carulli D, Manzo J, Coria-Avila GA (2016): Have we been ignoring the elephant in the room? Seven arguments for considering the cerebellum as part of addiction circuitry. Neurosci Biobehav Rev. 2016 Jan;60:1-11. doi: 10.1016/j.neubiorev.2015.11.005. PMID: 26602022. REVIEW ↥
Miquel M, Toledo R, García LI, Coria-Avila GA, Manzo J (2009): Why should we keep the cerebellum in mind when thinking about addiction? Curr Drug Abuse Rev. 2009 Jan;2(1):26-40. doi: 10.2174/1874473710902010026. PMID: 19630735. REVIEW ↥
Vazquez-Sanroman D, Carbo-Gas M, Leto K, Cerezo-Garcia M, Gil-Miravet I, Sanchis-Segura C, Carulli D, Rossi F, Miquel M (2015): Cocaine-induced plasticity in the cerebellum of sensitised mice. Psychopharmacology (Berl). 2015 Dec;232(24):4455-67. doi: 10.1007/s00213-015-4072-1. PMID: 26482898. ↥
Vazquez-Sanroman D, Leto K, Cerezo-Garcia M, Carbo-Gas M, Sanchis-Segura C, Carulli D, Rossi F, Miquel M (2015): The cerebellum on cocaine: plasticity and metaplasticity. Addict Biol. 2015 Sep;20(5):941-55. doi: 10.1111/adb.12223. PMID: 25619460. ↥
Miquel M, Gil-Miravet I, Guarque-Chabrera J (2020): The Cerebellum on Cocaine. Front Syst Neurosci. 2020 Oct 21;14:586574. doi: 10.3389/fnsys.2020.586574. PMID: 33192350; PMCID: PMC7641605. REVIEW ↥
Iversen L, Iversen F, Bloom S, Roth R (2009): Introduction to Neuropsychopharmacology, S. 181 ↥ ↥
Roy S, Field GD (2019): Dopaminergic modulation of retinal processing from starlight to sunlight. J Pharmacol Sci. 2019 May;140(1):86-93. doi: 10.1016/j.jphs.2019.03.006. PMID: 31109761. ↥
Prendergast BJ, Zucker I (2016): Ultradian rhythms in mammalian physiology and behavior. Curr Opin Neurobiol. 2016 Oct;40:150-154. doi: 10.1016/j.conb.2016.07.011. PMID: 27568859. REVIEW ↥
Goh GH, Maloney SK, Mark PJ, Blache D (2019): Episodic Ultradian Events-Ultradian Rhythms. Biology (Basel). 2019 Mar 14;8(1):15. doi: 10.3390/biology8010015. PMID: 30875767; PMCID: PMC6466064. REVIEW ↥
Yao T, Kim R (2025): Mathematical modeling of dopamine rhythms and timing of dopamine reuptake inhibitors. PLoS Comput Biol. 2025 Sep 25;21(9):e1013508. doi: 10.1371/journal.pcbi.1013508. PMID: 40997123; PMCID: PMC12494296. ↥ ↥ ↥ ↥ ↥
Oster H, Damerow S, Kiessling S, Jakubcakova V, Abraham D, Tian J, Hoffmann MW, Eichele G (2006): The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell Metab. 2006 Aug;4(2):163-73. doi: 10.1016/j.cmet.2006.07.002. PMID: 16890544. ↥
Korshunov KS, Blakemore LJ, Trombley PQ (2017): Dopamine: A Modulator of Circadian Rhythms in the Central Nervous System. Front Cell Neurosci. 2017 Apr 3;11:91. doi: 10.3389/fncel.2017.00091. PMID: 28420965; PMCID: PMC5376559. REVIEW ↥
Doyle SE, Grace MS, McIvor W, Menaker M (2002): Circadian rhythms of dopamine in mouse retina: the role of melatonin. Vis Neurosci. 2002 Sep-Oct;19(5):593-601. doi: 10.1017/s0952523802195058. PMID: 12507326. ↥
Chung S, Lee EJ, Yun S, Choe HK, Park SB, Son HJ, Kim KS, Dluzen DE, Lee I, Hwang O, Son GH, Kim K (2014): Impact of circadian nuclear receptor REV-ERBα on midbrain dopamine production and mood regulation. Cell. 2014 May 8;157(4):858-68. doi: 10.1016/j.cell.2014.03.039. PMID: 24813609. ↥
Colita CI, Hermann DM, Filfan M, Colita D, Doepnner TR, Tica O, Glavan D, Popa-Wagner A (2024): Optimizing Chronotherapy in Psychiatric Care: The Impact of Circadian Rhythms on Medication Timing and Efficacy. Clocks Sleep. 2024 Nov 5;6(4):635-655. doi: 10.3390/clockssleep6040043. PMID: 39584972; PMCID: PMC11586979. REVIEW ↥ ↥
Verwey M, Dhir S, Amir S (2016): Circadian influences on dopamine circuits of the brain: regulation of striatal rhythms of clock gene expression and implications for psychopathology and disease. F1000Res. 2016 Aug 24;5:F1000 Faculty Rev-2062. doi: 10.12688/f1000research.9180.1. PMID: 27635233; PMCID: PMC5007753. REVIEW ↥
Wesensten NJ, Belenky G, Kautz MA, Thorne DR, Reichardt RM, Balkin TJ (2002): Maintaining alertness and performance during sleep deprivation: modafinil versus caffeine. Psychopharmacology (Berl). 2002 Jan;159(3):238-47. doi: 10.1007/s002130100916. PMID: 11862356. ↥
McClung CA, Sidiropoulou K, Vitaterna M, Takahashi JS, White FJ, Cooper DC, Nestler EJ (2005): Regulation of dopaminergic transmission and cocaine reward by the Clock gene. Proc Natl Acad Sci U S A. 2005 Jun 28;102(26):9377-81. doi: 10.1073/pnas.0503584102. PMID: 15967985; PMCID: PMC1166621. ↥
McGeer EG, McGeer PL (1966): Circadian rhythm in pineal tyrosine hydroxylase. Science. 1966 Jul 1;153(3731):73-4. doi: 10.1126/science.153.3731.73. PMID: 5938420. ↥
Ferris MJ, España RA, Locke JL, Konstantopoulos JK, Rose JH, Chen R, Jones SR (2014): Dopamine transporters govern diurnal variation in extracellular dopamine tone. Proc Natl Acad Sci U S A. 2014 Jul 1;111(26):E2751-9. doi: 10.1073/pnas.1407935111. PMID: 24979798; PMCID: PMC4084435. ↥
Zawilska JB (1994): The role of dopamine in the regulation of melatonin biosynthesis in vertebrate retina. Acta Neurobiol Exp (Wars). 1994;54 Suppl:47-56. PMID: 7801792. REVIEW ↥
González S, Moreno-Delgado D, Moreno E, Pérez-Capote K, Franco R, Mallol J, Cortés A, Casadó V, Lluís C, Ortiz J, Ferré S, Canela E, McCormick PJ (2012): Circadian-related heteromerization of adrenergic and dopamine D₄ receptors modulates melatonin synthesis and release in the pineal gland. PLoS Biol. 2012;10(6):e1001347. doi: 10.1371/journal.pbio.1001347. PMID: 22723743; PMCID: PMC3378626. ↥
Sellix MT, Freeman ME (2003): Circadian rhythms of neuroendocrine dopaminergic neuronal activity in ovariectomized rats. Neuroendocrinology. 2003 Jan;77(1):59-70. doi: 10.1159/000068334. PMID: 12624542. ↥
Hampp G, Ripperger JA, Houben T, Schmutz I, Blex C, Perreau-Lenz S, Brunk I, Spanagel R, Ahnert-Hilger G, Meijer JH, Albrecht U (2008): Regulation of monoamine oxidase A by circadian-clock components implies clock influence on mood. Curr Biol. 2008 May 6;18(9):678-83. doi: 10.1016/j.cub.2008.04.012. PMID: 18439826. ↥
Taufique SKT, Ehichioya DE, Pendergast JS, Yamazaki S (2022): Genetics and functional significance of the understudied methamphetamine sensitive circadian oscillator (MASCO). F1000Res. 2022 Sep 8;11:1018. doi: 10.12688/f1000research.125432.2. PMID: 36226037; PMCID: PMC9539084. ↥
Abarca C, Albrecht U, Spanagel R (2002): Cocaine sensitization and reward are under the influence of circadian genes and rhythm. Proc Natl Acad Sci U S A. 2002 Jun 25;99(13):9026-30. doi: 10.1073/pnas.142039099. PMID: 12084940; PMCID: PMC124417. ↥