Liebe Leserinnen und Leser von ADxS.org, bitte verzeihen Sie die Störung.
ADxS.org benötigt in 2024 rund 58.500 €. Leider spenden 99,8 % unserer Leser nicht.
Wenn alle, die diese Bitte lesen, einen kleinen Beitrag leisten, wäre unsere Spendenkampagne für das Jahr 2024 nach wenigen Tagen vorbei. Dieser Spendenaufruf wird 23.000 Mal in der Woche angezeigt, jedoch nur 75 Menschen spenden.
Wenn Sie ADxS.org nützlich finden, nehmen Sie sich bitte eine Minute Zeit und unterstützen Sie ADxS.org mit Ihrer Spende. Vielen Dank!
Seit dem 01.06.2021 wird ADxS.org durch den gemeinnützigen ADxS e.V. getragen. Spenden an den ADxS e.V. sind steuerlich absetzbar (bis 300 € genügt der Überweisungsträger als Spendenquittung).
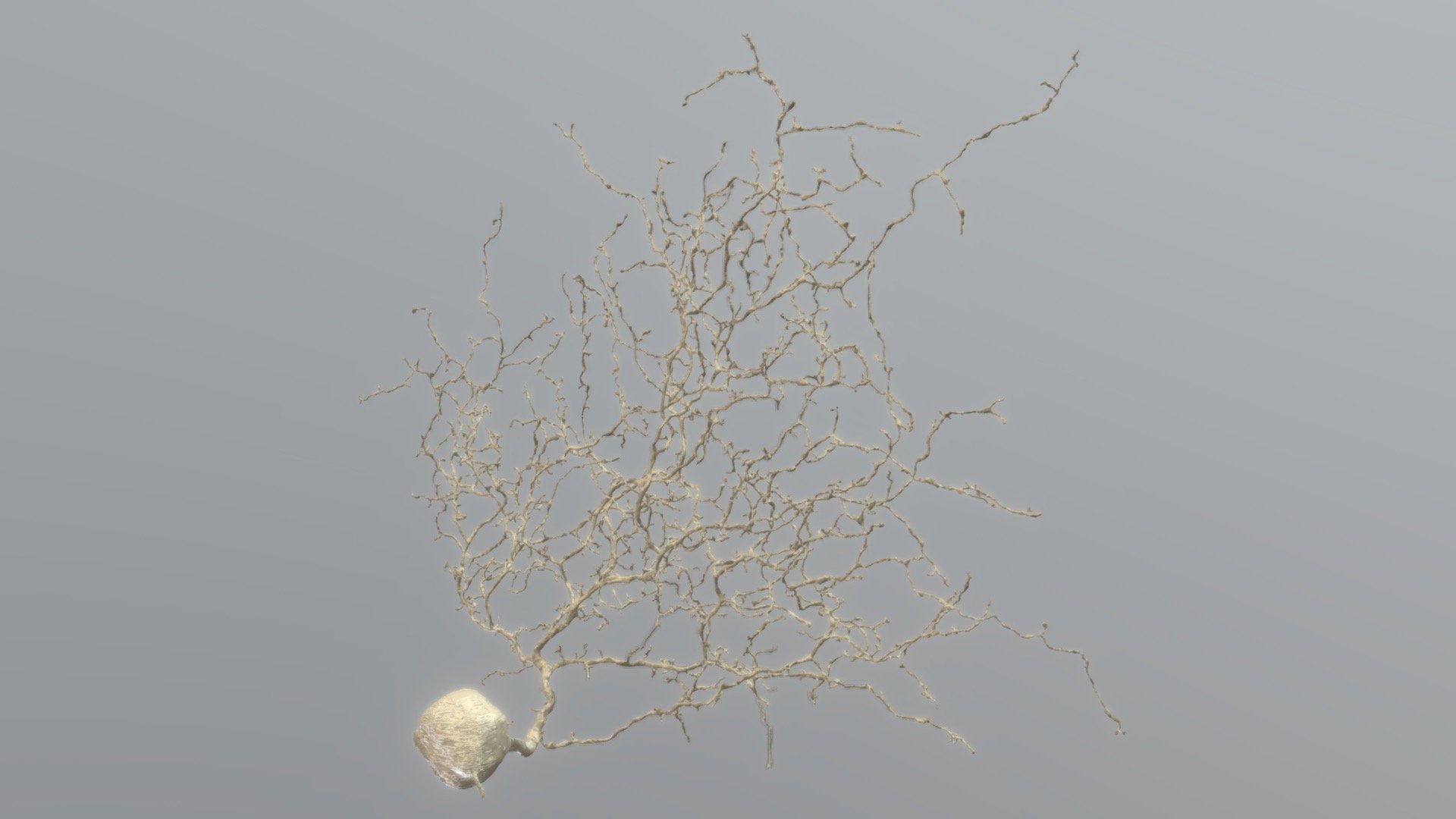
Zytoplasma (bestehend aus dem Zytosol, der Zellflüssigkeit, und den darin schwimmenden Organellen (Mitochondrien, Golgi etc.))
dem Nukleus (dem Zellkern im Soma)
vielen Dendriten (an denen sie durch zigtausende bis Millionen Synapsen Informationen von anderen Zellen empfangen) und
einem Axon (Neurit), über das sie Informationen an andere Zellen senden.
Während Dendriten maximal einige hundert Mikrometer kurz sind, können Axone beim Menschen zwischen 0,1 Millimeter und über einem Meter (oder sogar bis zu 4 Meter1] lang sein. Das Axoplasma innerhalb des Axons umfasst mehr als 90 % des Zytosols. Der Axonhügel ist der Ursprung des elektrischen Signals der Nervenzelle und verbindet Zellkern und Axon. Am Ende eines Axons befinden sich Terminale, die die Information der präsynaptischen Zelle über chemische Synapsen an postsynaptische Zellen weitergeben. Die axonalen Synapsen docken meist an Dendriten, zuweilen an Soma und selten auch an Axone anderer Zellen an.
Axone können eine Myelinscheide aus Schwannzellen haben. Diese “isoliert” die elektrische Leitung des Axons und bewirkt mittels der Schnürringe, an denen die Isolierung unterbrochen ist, einer regelmäßige Verstärkung des weiterzuleitenden elektrischen Signals. Je dicker das Axon, und je besser es von Gliazellen (im Gehirn Oligodentrozyten, peripher Schwannzellen) ummantelt (myelinisiert) ist, desto schneller ist die elektrische Weiterleitung (bis zu 120 Meter / Sekunde).
Docken genug Neurotransmitter an die exzitatorischen Rezeptoren eines Neurons und wenig genug an inhibitorische Rezeptoren, wird ein Aktionspotenzial ausgelöst. Das Neuron feuert dieses als elektrischen Impuls über das Axon zu den aktiven Zonen an den Synapsen, wo dadurch Vesikel mit der Zellmembran verschmelzen und Neurotransmitter in den synaptischen Spalt freisetzen.
Nervenzellen können bis zu 500-mal/Sekunde feuern.
Das menschliche Gehirn hat ca. 100 Milliarden Nervenzellen.
Davon sind:2
glutamaterg - ca. 20.000.000.000 (20 Milliarden) GABAerg - ca. 9.000.000.000 (8 Milliarden)
serotonerg - ca. 250.000 dopaminerg - ca. 250.0002, 400.000 bis 600.000 noradrenerg - ca. 30.000 bis 50.000
Informationen werden durch eine gleichzeitige Feuerung einer jeweiligen Gruppe von Nervenzellen repräsentiert.
Unipolare, bipolare und multipolare Neuronen
Bei unipolaren Neuronen ist das Axon ein Zweig des Dendriten. Unipolare Neuronen finden sich vor allem in Nervensystemen von Nichtwirbeltieren vor sowie im autonomen Nervensystem von Wirbeltieren.
Bipolare Neuronen haben ein ovales Soma, aus dem auf der einen Seite der Dendritenbaum und auf der anderen das Axon entspringen. Sinneszellen sind meist bipolare Neuronen.
Bei multipolaren Neuronen entspringen dem Soma eine Vielzahl von Dendriten sowie ein Axon. Die meisten Nervenzellen des Gehirns sind multipolar.
Sinnesneurone, Motorneurone, Interneurone
Sinnesneurone empfangen Sinnessignale (Druck, Temperatur, Licht etc.) aus der Peripherie und senden diese an das Rückenmark.
Motorneurone leiten Signale vom Gehirn und Rückenmark an Muskeln und Drüsen. Interneurone verbinden zwei andere Neurone untereinander (was die meisten Neuronen tun).
Lokale Interneurone verbinden nahegelegene Neurone und haben daher kurze Axone; Relay- oder Projektions-Interneurone leiten das Signal in andere Gehirnregionen und haben daher lange Axone.
Alle Zellen im Nervensystem, die keine Neuronen sind, heißen Gliazellen.
Im Nervensystem von Wirbeltieren finden sich 2- bis 10-mal so viele Gliazellen wie Neuronen. Gliazellen senden keine Signale, sondern unterstützen Nervenzellen.
Astrozyten ernähren die Neuronen über Kontakte zu Blutgefäßen. Astrozyten sind maßgeblich an der Flüssigkeitsregulation im Gehirn beteiligt und sorgen für die Aufrechterhaltung des Kalium-Haushaltes.
Astrozyten (Astroglia) ist der Namen dieser Zellen im zentralen Nervensystem. (Gehirn / Rückenmark)
Mantelzellen (Sateliltenzellen) heißen diese Zellen, wenn sie im Körper (peripher) vorkommen.
Mikroglia sind Immuneffektorzellen im ZNS. Sie zählen lediglich formal zur Familie der Gliazellen. Genauer sind es Zellen des mononukleär-phagozytären Systems.
Synapsen sind die Verbindungsstellen, über die Nervenzellen miteinander kommunizieren.
Es gibt
elektrische Synapsen, die elektrische Signale direkt und damit sehr schnell weitergeben, was jedoch nur an kleinere Neuronen möglich ist und
chemische Synapsen, bei denen das Signal über Neurotransmitter vermitteln, was erstens eine modulierende Steuerung ermöglicht und zweitens eine Verstärkung des Signals, sodass auch größere Neuronen adressiert werden können.
Während Muskelzellen meist nur von einem einzigen Motorneuron exzitatorisch gesteuert werden, und jedes einzelne Signal eine Muskelaktivierung bewirkt, sind Neuronen im Gehirn vielfach und redundant miteinander vernetzt, können anregend (exzitatorisch) oder hemmend (inhibitorisch) miteinander verbunden sein und benötigen für eine Aktivierung ein Zusammenwirken von vielen Signalen (oft 50 bis 100).
Neurotransmitter sind Botenstoffe, die Informationen an chemischen Synapsen zwischen Nerven übertragen. Beispiele sind Dopamin, Noradrenalin, Serotonin, Acetylcholin, GABA und Glutamat. Die unterschiedlichen Neurotransmitter haben verschiedene Aufgaben im Gehirn und überlappen sich in ihrer Wirkung. Neurotransmitter bewirken durch ihre Ausschüttung an den Synapsen eine chemische Reizweiterleitung bzw. -blockade zwischen Neuronen (Nervenzellen).
Andere Botenstoffe, die Hormone, vermitteln ihre Wirkung langsam über die Blutbahn an weiter entfernte Zielorgane (z.B. Adrenalin, Cortisol, Estradiol, Insulin, Testosteron, Thyroxin, Triiodthyronin).
Manche Stoffe wirken zugleich als Neurotransmitter wie als Hormone (z.B. Noradrenalin, Serotonin, Histamin).Manche Stoffe wirken zugleich als Neurotransmitter als auch als Hormone (z.B. Noradrenalin, Serotonin, Histamin).
Neurotransmitter werden im Cytosol des Zellkerns synthetisiert, in Vesikel verpackt und über die Mikrotubuli durch die Axone zu den Nerventerminalen transportiert, wo sich die sendenden Synapsen befinden. Die Transportgeschwindigkeit in den Axonen ist je nach Substanz unterschiedlich und beträgt bis zu 5 µm/Sekunde = ca. 40 cm / Tag.3
Aus den Nerventerminalen werden die Neurotransmitter auf ein Aktionspotenzial in den synaptischen Spalt ausgeschüttet, indem sich die Vesikel mit der Membran verbinden. Der synaptische Spalt ist zwischen 20 und 40 nm breit.4
Im synaptischen Spalt docken sie an postsynaptische Rezeptoren an und öffnen dadurch Ionenkanäle, was in der empfangenden Zelle ein neues Aktionspotential auslösen kann.
Danach lösen sich die Neurotransmitter wieder von den postsynaptischen Rezeptoren und werden im oder am Rande des synaptischen Spalts durch präsynaptische Transporter in die sendende Zelle wiederaufgenommen. In der Zelle werden sie entweder erneut in Vesikel eingelagert, bis zur nächsten Ausschüttung, oder durch abbauende Enzyme verstoffwechselt (z.B. Dopamin und Noradrenalin durch Monoaminoxidase und COMT).
Nicht wiederaufgenommene Neurotransmitter diffundieren in den extrazellulären Raum und können von dort aus D2-Autorezeptoren der sendenden Zelle oder Rezeptoren oder Transporter anderer Zellen ansteuern.
Zweck der chemischen Signalübertragung durch Neurotransmittern ist die Filterung und Modulation von Signalen und die Möglichkeit einer Verstärkung des zu übertragenden Signals.
Inverted-U: Zu hohe wie zu niedrige Neurotransmitterspiegel beeinträchtigten Informationsübertragung bei Katecholaminen
Eine optimale Informationsübertragung erfordert - jedenfalls bei Katecholaminen (wie Dopamin, Noradrenalin, Adrenalin) - einen optimalen Neurotransmitterspiegel. Ein verringerter wie ein erhöhter Spiegel beeinträchtigen die Signalübertragung. Eine optimale Informationsübertragung zwischen Gehirnsynapsen erfordert einen optimalen Pegel der beteiligten Neurotransmitter. Ein zu geringer Neurotransmitterspiegel führt zu nahezu identischen Folgen der Signalübertragungsstörung wie ein zu hoher Neurotransmitterspiegel (Inverted-U-Theorie).56
Die Aktivität des Locus coeruleus (Hauptquelle für kortikales Noradrenalin) zeigt eine Inverted-U-Relation zur Aufgabenleistung7
Die DRD1-Aktivierung steht in einer Inverted-U-Relation zur Leistung des Arbeitsgedächtnisses89
Die Aktivität der DA-Neuronen im VTA steht in einer Inverted-U-Relation zur Leistung des Kurzzeitgedächtnisses10
Es gibt rund 500 verschiedene Neurotransmitter in verschiedenen Neurotransmitterklassen.
Lösliche Gase
Stickstoffmonoxid (NO)
Kohlenstoffmonoxid (CO)
Schwefelwasserstoff
Amine
Choline (quartäre Amine)
Acetylcholin
Biogene Amine
(Klassische) Monoamine
Katecholamine:
Noradrenalin
Adrenalin
Dopamin
Indolamine
Serotonin
Melatonin
Imidazolamine
Histamin
Spurenamine
Phenethylamine
Phenethylamin (PEA)
Tyramin
Indolamine
Tryptamin
Octopamin
Aminosäuren
Inhibitorische Aminosäuretransmitter
Gamma-Amino-Buttersäure (GABA)
Glycin
β-Alanin
Taurin
Exzitatorische Aminosäuretransmitter
Glutaminsäure (Glutamat)
Asparaginsäure (Aspartat)
Cystein
Homocystein
Neuropeptide
Opioipeptide
Dynorphine
Dynorphin A
Dynorhin B
α-Neoendorphin
β-Neoendorphin
Endorphine
Somatostatin
Insulin
Glucagon
α-Endopsychosin
Neurokinine / Tachykinine
Substanz P (Neurokinin 1)
Neurokinin A (Substanz K)
Neuropeptid K (Neurokinin K)
Neuropeptid γ (Neuropeptid gamma)
Neurokinin B
Hemokinin-1
Endokinin A, B, C und D
Enkephaline
Met-Enkephalin
Leu-Enkephalin
Met-Arg-Phe-Enkephalin
sonstige Neuropeptide
Oxytocin
Somatostatin
Vasopressin
Neuropeptid S
GHRH
Endocannabinoide
Anandamid
2-Arachidonylglycerol
O-Arachidonylethanolamid
Neurotransmitter geben je nach Rezeptor, an den sie andocken, aktivierende oder inhibierende (hemmende) Informationen weiter. Dopamin und Serotonin sind zwar überwiegend bei der Weitergabe inhibierender Informationen beteiligt, doch sind nur die D2, D3 und D4 Rezeptoren inhibierend (sie hemmen das Enzym Adenylylcyclase), während die D1 und D5 Rezeptoren aktivierend (exzitatorisch) wirken (sie aktivieren das Enzym Adenylylcyclase).
Bei ADHS ist die Informationsübertragung im Gehirn vornehmlich in Bezug auf die Neurotransmitter Dopamin und Noradrenalin beeinträchtigt.
Rezeptoren sind Andockstellen für Botenstoffe. Je nach Rezeptor kann ein Botenstoff hemmend oder aktivierend wirken.
Ionotrope Rezeptoren
Bei diesen bewirkt die Bindung eines Neurotransmitters unmittelbar die Öffnung eines Ionenkanals.
Metabotrope Rezeptoren
Bei diesen bewirkt die Bindung eines Neurotransmitters eine Aktivierung von Botenstoffen (Second Messenger), die wiederum etliche Transportkanäle adressieren können.
Alle Dopamin- und Noradrenalinrezeptoren sind metabotrop.
Exzitatorische und inhibitorische Rezeptoren.
Rezeptoren für ein und denselben Neurotransmitter können exzitatorisch (die postsynaptische Nervenspannung erhöhend) und inhibitorisch (die postsynaptische Nervenspannung verringernd) wirken.
Dagegen sagen weder das Feuern von Neuronen noch die Art des Neurotransmitters etwas darüber aus, ob ein Signal aktivierend (exzitatorisch) oder hemmend (inhibitorisch) sein soll.
Neuronen vermitteln Signale, indem sie ein Aktionspotential auslösen. Neuronen enthalten im Ruhezustand in ihrem Inneren eine um durchschnittlich 65 mV (je nach Zellart zwischen 45 und 90 mV) niedrigere Spannung als der extrazelluläre Raum. Dieser Spannungsunterschied entsteht, indem die sogenannte Natrium-Kalium-Pumpe (Natrium-Kalium-ATPase, ein Membranprotein), Natriumionen aus dem Zellinneren gegen Kaliumionen aus dem Extrazellulärraum tauscht. Durch kaliumdurchlässige Ionenkanäle in der selbst undurchlässigen Zellmembran können die Kaliumionen - dem Konzentrationsgefälle folgend - langsam wieder die Zelle verlassen und hinterlassen dabei eine nicht neutralisierte negative Ladung an der inneren Zellmembranoberfläche, die in der Regel bei -65 mV liegt.
In den Zellen befinden sich dann im Ruhezustand rund 1/10 an Natriumionen und das 20-fache an Kaliumionen wie extrazellulär. Das extrazelluläre Natrium und Kaliumionenniveau wird durch die Nieren und die Astrozyten aufrechterhalten. Gelangen Natrium- oder Calciumionen in die Zelle, steigt deren Spannung.
Ein Aktionspotential (ein schneller Spannungsanstieg um +10 mV, z.B. von -65 auf -55 mV) macht die Zellmembran durchlässiger für Natriumionen als für Kaliumionen. Der dadurch gesteigerte Eintritt von Natriumionen erhöht die Zellwanddurchlässigkeit für Natriumionen weiter, sodass immer mehr Natriumionen eintreten. Dadurch verringert sich die negative Spannung schlagartig und verkehrt sich sogar kurzfristig (für rund 1 ms) ins Positive auf + 40 mV (“Overshoot”). Das Aktionspotential wandert nun mit 1 bis 100 Metern/Sekunde das Axon entlang bis zu den Terminalen, wo es Ionenkanäle öffnet.
Das Aktionspotential ist eine Alles-oder-Nichts-Entscheidung. Wird es ausgelöst, hat es stets die volle Stärke, egal ob die Auslösungsschwelle nur ganz knapp oder stark überschritten ist.
Das Aktionspotential bleibt über die gesamte Strecke im Axon konstant. Dazu wird es an den Ranvier-Schnürringen verstärkt.
Nachdem das Spannungsmaximum erreicht ist, erfolgt durch das Schließen von Natrium- und das Öffnen von Kaliumkanälen die Rückkehr zum Ruhepotential (Repolarisation).
Dabei wird die Membranspannung zunächst noch negativer, als das ursprüngliche Ruhepotential war (Hyperpolarisation). Danach kehrt die Zelle zum Ausgangspunkt zurück (Ruhepotential).
Nach der Auslösung eines Aktionspotentials unterliegt ein Neuron einer Pause, der Refraktionsperiode.
Spannung steigt (langsam oder schnell) in Richtung Schwellenpotenzial, z.B. von -70 mV auf -50 mV (initiale Depolarisation)
Erreichen eingehende Reize (nach Summierung im Axonhügel) den Schwellwert nicht, bleibt es bei einer vorübergehenden, reversible Veränderung des Membranpotentials
Aufstrich
Nur und erst bei Erreichen des Schwellenpotenzials erfolgt eine vollständige Depolarisation
Folge:
Die spannungsabhängigen Natriumkanäle öffnen sich und lassen aus dem Extrazellulärraum schlagartig Na+-Ionen in das Zytosol des Neurons einströmen
Die Kaliumkanäle sind währenddessen geschlossen
Ein positiver Rückkopplungsmechanismus bewirkt am Ende gar eine Ladungsumkehr (“Overshoot”).
Repolarisation
Natriumkanäle beginnen noch vor dem Potentialmaximum, sich wieder zu schließen
Die spannungsabhängigen Kaliumkanäle öffnen sich, sodass K+-Ionen aus dem Zellinnerem in den Extrazellulärraum strömen
Die Leitfähigkeit der Kaliumkanäle erreicht ihr Maximum, wenn fast alle Natriumkanäle bereits inaktiviert sind
Während der Repolarisation bewegt sich das Potential wieder in Richtung des Ruhepotentials, was zur Schließung der Kaliumkanäle führt, während die Natriumkanäle langsam wieder aktiviert werden.
Hyperpolarisation
Kaliumkanäle schließen sich binnen 1 bis 2 ms, und damit langsamer als Natriumkanäle
Währenddessen sinkt das Membranpotential unter das eigentliche Ruhepotential (“Hyperpolarisation”)
Refraktärzeit
nach einem Aktionspotential ist das Neuron für eine kurze Zeit nicht erregbar
so lange, bis die Natriumkanäle wieder aktivierbar sind
Absolute Refraktärphase: Zeitspanne kurz nach dem Overshoot, vor Abschluss der Repolarisation. Aktionspotential nicht auslösbar.
Relativen Refraktärphase: Schwellenwert für die Auslösung eines Aktionspotentials ist erhöht
Während das Aktionspotential immer gleich stark und der einzige ausgehende Impuls eines Neurons ist, gibt es zwei Arten von aktivierenden Impulsen:
das synaptische Signal
das Rezeptorsignal.
Beide sind von der Stärke her abgestuft (graduell).
Rezeptorsignale werden beispielsweise durch periphere sensorische Reize ausgelöst. Ein Rezeptorsignal entspricht in Dauer und Stärke der Intensität des Reizes, ist aber insgesamt relativ schwach. Es reicht innerhalb des Neurons nur wenige Millimeter weit. Nach einem Millimeter hat es bereits zwei Drittel seiner Energie verloren. Erreicht es innerhalb seiner Reichweite mit ausreichender Stärke einen Ranvier-Schnürring, wird durch dessen Verstärkungseffekt ein vollständiges Aktionspotential ausgelöst, sodass der sensorische Reiz das Rückenmark erreichen kann.
Synaptische Signale werden durch Neurotransmitterbindung an empfangenden Synapsen an Dendriten ausgelöst. Sie sind, wie das Rezeptorsignal, graduell je nach Menge der aktivierten Rezeptoren. Synaptische Signale werden am Axonhügel des Neurons aufsummiert. Wenn die Summe den Schwellwert überschreitet, wird das Aktionspotential ausgelöst.
Auch wenn ein Aktionspotential immer gleich stark ist, kann es durch die Häufigkeit und Frequenz seiner Abfolge eine graduell abgestufte Neurotransmitterausschüttung bewirken und so wieder unterschiedlich starke Signale an die postsynaptisch verbundenen Zellen weitergeben. Wird ein Aktionspotential nur einmal ausgelöst. Je häufiger und schneller hintereinander das Aktionspotential ausgelöst wird, desto größer ist die Menge der Transmitterausschüttung, was zu einer höheren Anzahl an postsynaptisch adressierten Rezeptoren führt.
Das menschliche Gehirn wird von rund 600 km Blutgefäßen durchzogen.
Blutgefäße im Gehirn verfügen in ihren Wänden über besondere Zellen, die verhindern, dass bestimmte Stoffe, die im Körper (peripher) unschädlich sind, die komplexen und empfindlichen Abläufe des Gehirns (zentral) stören, und dass Neurotransmitter und Kalium aus der extrazellulären Gehirnflüssigkeit ins Blut austreten.
Nur fettlösliche Substanzen mit einem Molekulargewicht unter 500 Da können durch die Blut-Hirn-Schranke diffundieren, wie z.B. Nikotin, Alkohol, Blutgase oder Narkotika wie Halothan, nicht aber Ionen oder polare Substanzen wie Glucose. Letztere sind auf spezifische Transportsysteme angewiesen.12
Im Laufe der Evolution haben sich nach und nach verschiedene Gehirnbereiche entwickelt, die wir nachfolgend von alt nach neu darstellen.
Grundlegend zu den Gehirnbereichen des Menschen: Kandel, Shadlen (2021): The Brain and Behaviour. In: Kandel, Koester, Mack, Siegelbaum (2021): Principles of Neuronal Science.
jede Hemisphäre ist in 4 Großhirnrindenlappen geteilt
frontal (PFC)
höhere kognitive Prozesse
Kontrolle der Willkürmotorik, Aufmerksamkeit, Kurzzeitgedächtnisaufgaben, Motivation und Planung
parietal (Parietallappen, Scheitellappen, oben)
somatosensorische Funktionen
visuelle Steuerung von Bewegungen und Erkennung von Reizen im Raum
räumliches Denken und „quasi-räumliche“ Prozesse wie Rechnen und Lesen
Sprachverarbeitung
Schnittstelle zwischen den Sinnessystemen (vor allem des visuellen Systems) und dem motorischen System für die Berechnung, Ausführung und Kontrolle von Hand und Augenbewegungen.
okzipital (hinten)
visueller Cortex (visuelle Verarbeitung)
temporal (Seitenlappen)
auditorischer Cortex (akustische Verarbeitung)
Deutung der Informationen, entsprechend des visuellen Gedächtnisses und des Sprachverständnisses
9. Gehirnregionen und Funktionen - Hardware und Software¶
Zwar haben einzelne Gehirnregionen bevorzugte Funktionen, jedoch besteht keine eindeutige 1:1 Zuordnung.
Einfache Reflexe werden noch recht eindeutig durch bestimmte Gehirnregionen gesteuert. Je komplexer eine Verhaltensfunktion ist, desto mehr wird das Zusammenspiel mehrerer Gehirnregionen genutzt - hier spricht man von Gehirnnetzwerken.
Es hilft einem Individuum beim Überleben, wenn wichtige Funktionen alternativ von verschiedenen Gehirnregionen gesteuert werden können (Redundanz). Ausfallende Funktionen (z.B. wenn eine Gehirnregion durch einen Schlaganfall geschädigt wird) können dadurch auch durch andere Gehirnregionen übernommen werden (Flexibilität).
Ohne die Übernahme der Steuerung der beeinträchtigten Funktion durch eine andere Gehirnregion wäre es zwar möglich, die Verschlechterung oder den Totalausfall einer Funktion durch entsprechende Verlagerung der Überlebensstrategien auf eine verstärkte Nutzung anderer Fähigkeiten (Verhaltensänderung) auszugleichen. Es ist jedoch wesentlich einfacher, eine bereits als erfolgreich erlerntes Verhalten beizubehalten, indem wichtige Funktionen aufrechterhalten werden können, weil sie durch mehr als lediglich eine Gehirnregion repräsentiert werden und lediglich die Ansteuerung der Funktion neu erlernt werden muss.
Dies erklärt zugleich die Schwierigkeiten, bestimmte Funktionsbeeinträchtigungen auf Defekte bestimmter Gehirnregionen zurückzuführen. Dies betritt insbesondere die Mechanismen, die keine spezifischen Körperfunktion repräsentieren.
Die bei ADHS bestehenden Probleme betreffen eher Mechanismen zur langfristigeren Regulation von Verhaltensweisen und können nicht bestimmten Hardware-Defekten einzelner Gehirnregionen zugeordnet werden.

{kind=link}
-1.svg){kind=link}